




1)腎臓発生の概略
腎臓は中間中胚葉から発生し、前腎、中腎、後腎の3段階を経て形成される。前腎、中腎のほとんどは後に退行変性し、哺乳類成体において機能する腎臓は後腎である。(図1)に示すように、前腎は1つのネフロンからなる非常に単純な構造である。中腎はその尾側に発生し、数十のネフロンからなる。この一部は男性生殖器となるが、腎臓としての中腎は退行する。我々哺乳類の腎臓、すなわち後腎は、中腎管 (ウルフ管)の最も尾側に尿管芽と呼ばれる突起が出現し、その周りに間葉組織が集合して生じる。この尿管芽と後腎間葉との相互作用によって、数百万ものネフロンをもつ後腎が完成する。尿管芽の周りに間葉細胞が凝集し、それが上皮化してS字体といわれる状態を経て、糸球体、近位及び遠位尿細管が発生する(図2)。尿管芽は分岐を重ね、集合管、尿管となる。つまり、腎臓としての機能を司るそのかなりの部分が後腎間葉から発生することになり、尿細管には10種を超える細胞が存在するため、後腎間葉は多能性をもった前駆細胞集団ともいえる。
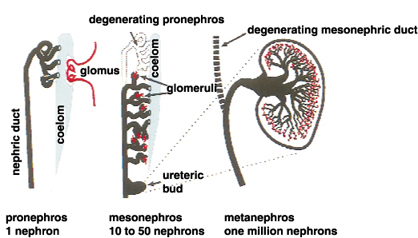
図1 腎臓の構造
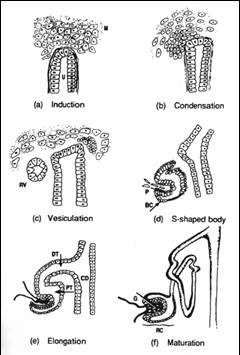
図2 ネフロンの発生
2)Znフィンガー蛋白 Sall1の単離
前腎はアフリカツメガエル(Xenopus laevis)で解析が進んでおり、特にアニマルキャップアッセイという予定外胚葉を培養して各種臓器を誘導する系が確立している。東京大学の浅島誠らによって、このアニマルキャップをアクチビンとレチノイン酸の存在下に生理食塩水の中で培養すると、わずか3日で3次元の立体構造をもった前腎管が形成されることが示されている。我々はこの系に着目し、浅島らと共同して、この前腎管を誘導する条件としない条件とで、種々のサブトラクションを行った(図3A)。その中から単離されたのが、Zincフィンガーを8個持つ新規蛋白で、これをXsal-3と名付けた(図3B)。この遺伝子はショウジョウバエのspalt (sal)のホモログで、確かに前腎に発現していた。この遺伝子を指標にマウス後腎から新たな遺伝子が単離でき、この遺伝子(Sall1)の発現様式を調べたところ、尿管芽が後腎間葉に侵入する胎生11.5日において、尿管芽を取り囲む後腎間葉に非常に強く発現していた(図4)。Sall1は腎臓の他に、中枢神経系、耳胞、心臓、肢芽、肛門などに発現しており、Xsal-3との一部類似性が認められた。さらに、中枢神経系では脳室周囲の神経幹細胞が存在する領域に、肢芽ではprogress zoneという未分化細胞が増殖する部分で発現が認められ、腎臓に限らず他の未分化細胞でも何らかの役割を持つ可能性が示唆された。
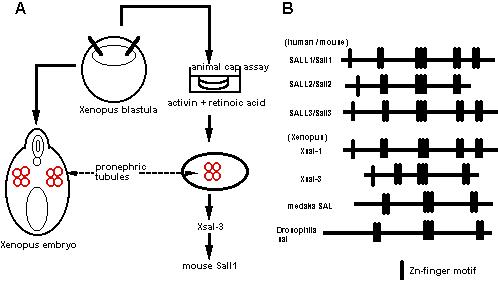
図3 Salの単離
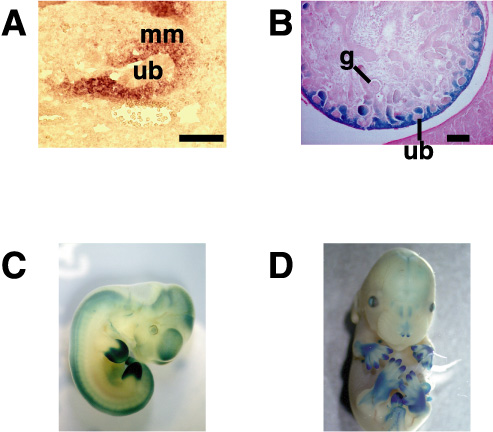
図4 マウスSall1の発現パターン
3)Sall1は腎臓発生に必須である
そこでSall1を欠失するマウスを作成したところ、すべてのノックアウトマウスが生直後に死亡した。開腹してみると、腎臓が完全に欠損しているか、非常に小さい痕跡的な腎臓が認められるのみであったため(図5)、Sall1は腎臓の発生に必須であることが証明された。さらにノックアウトマウスでは、尿管芽は後腎間葉に侵入していないか、あるいはしてもその後の分岐は著明に障害されていた。つまりSall1が、後腎発生の最も初期段階の重要なステップである尿管芽の侵入に必須であることが判明した。
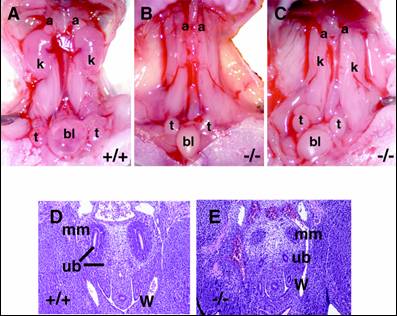
図5 Sall1ノックアウトマウス
後腎間葉は尿管芽の分岐とともにそれを取り囲むように分布するので、どんどん数が増え一つ一つは小さくなっていく。齧歯類においてはこのような間葉組織は生後数日まで存在するが、成体においては認められない。この間葉組織が失われることが、成体において腎臓が修復しない原因ではないだろうか?とすれば、腎臓の前駆細胞集団である間葉組織を何らかの方法で誘導したり移植したりできれば成体での腎臓修復も可能になるかもしれない。これを実現するためには腎臓前駆細胞のアッセイ系が必須である。そこで我々は発生期腎臓から前駆細胞を同定する系を確立した。Sall1は我々が単離した核内因子で、そのノックアウトマウスは腎臓を欠損する。この遺伝子座にGFPを導入したマウスでは、腎臓前駆細胞集団である後腎間葉が蛍光発色する。GFPが高発現する細胞を FACS で選別し、Wnt4を発現するフィーダー上で培養すると、1個の細胞からコロニーが形成され、このコロニーは糸球体、近位尿細管、遠位尿細管という多系統へ分化することを見いだした。さらにGFP高発現の細胞群を再凝集させ器官培養すると3次元構造を再構築できることを示した。これは腎臓前駆細胞をprospectiveに同定する初めての系であり、組織や切片のレベルでしか検討されていなかった腎臓形成が、単一細胞レベルで解析できるようになる可能性を秘めている。またこのコロニーアッセイを指標として、胚性幹(ES)細胞から腎臓前駆細胞を誘導できるかもしれない。これらの結果はDevelopment誌に発表され、巻頭でも紹介された。
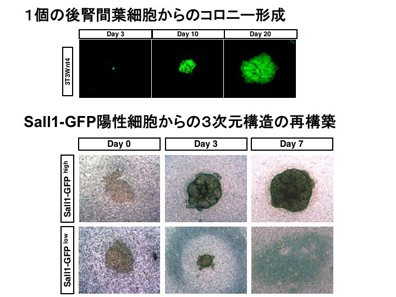
図6 当研究室で開発した腎間葉細胞の培養系
我々はこれまでに核内因子 Sall1が腎臓発生に必須であることを示してきた。さらに、そのファミリーであるSall4のノックアウトマウスを作成した。Sall4を欠失する胚は子宮着床直後に死亡し、胎児のおおもとである内部細胞塊の増殖が障害されていた。この内部細胞塊由来の細胞株が、再生医療で注目されている胚性幹(ES)細胞である。そこでES細胞でSall4を欠失させたところ増殖が著明に低下し、Sall4 がES細胞に必須であるという予想外の事実が明らかになった。一方Sall4のヘテロマウスは肛門と心臓の異常を呈し、ヒトSall4の変異でおきるOkihiro症候群の症状の一部を再現できた。さらに、共にヒト疾患の原因である Sall4とSall1 が二量体を形成して臓器形成に関わること、ヒト Sall1 の変異によって生じる症状の一部は、変異型 Sall1 が Sall4 の機能を阻害するためであることを明らかにした。この研究成果は Development 誌 8 月号に発表され、巻頭でも“Cooperative SALLies form organs(Sallが協調して器官を作る、Cooperative SALLiesには共同戦隊の意味もひっかけてある)”として、紹介された。Sallファミリーが腎臓に限らず様々な臓器、さらにはES細胞にも必須であることが判明し、その分子機構解明がさらに重要性を帯びてきた。
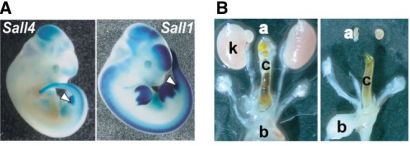
図 A.Sall4とSall1の発現は一部重なっている(矢頭は肛門)
B.Sall4/Sall1の二重ヘテロマウス(右)は腎臓を欠損する。左は正常マウス。
a:副腎、b:膀胱、c:腸、k:腎臓
Sall1 強陽性の腎臓の前駆細胞で発現するキネシンKif26bを同定し、そのノックアウトマウスを作成したところ、腎臓が完全に欠損し、 Sall1 の場合と同様に尿管芽の侵入が障害されていた。 Sall1の直接の下流であり、かつその機能のかなりの部分を説明できる因子である。Kif26b はミオシンを介して後腎間葉細胞の接着を制御し、インテグリンα 8 さらには GDNF の発現を維持していることが示唆された。単一のキネシンの欠失によって臓器が丸ごと欠損するという報告は今回が初めてである。本研究は、遺伝子断片の単離から7年を費やした 労作であり、 Proc. Natl. Acad. Sci. USA 誌電子版( 2010 年 5 月)に先行掲載された。→発生医学研究所ホームページ「New Press」での紹介記事
腎臓発生分野では、カエルとマウスを用いて腎臓形成に関わる遺伝子群を同定してきました。 タンパク質脱リン化酵素 Dullard も発生期のカエルの腎臓から単離されたもので、 BMP シグナルを抑制することによってカエルの初期発生を制御します。しかし遺伝子の同定から 10 年以上過ぎても、 Dullard の哺乳類における役割は腎臓を含めて不明のままでした。そこで腎臓の前駆細胞で Dullard を欠損させたマウスを作成したところ、予想に反して胎児期の腎臓はほぼ正常に形成されました。しかし出生後数週内に腎臓の中心部が細胞死によって消失して空洞化し、すべてのマウスが成体になる前に死亡しました。 Dullard が欠損することによって、 BMP シグナルが過剰に入って腎臓の細胞死が引き起こされたためでした。さらにこの Dullard 欠損マウスに BMP シグナルの阻害薬 LDN193189 を投与すると、腎臓の症状が改善されました。よって Dullard が BMP シグナルを抑制して適度なレベルに保つことが、出生後の腎臓の維持に必須であることが明らかになりました。この研究は、今まで見過ごされがちだった出生後つまり小児期の腎臓の維持の重要性を示すもので、小児期の腎臓疾患の病因解明や治療に向けた重要な情報となります 。 Dullard や BMP シグナルの異常がヒトの病気の原因になっているか、それを BMP シグナルの阻害薬で 治療できるか、など 今後の展開が期待されます。この研究成果は 2013 年 1 月 29 日に Nature Communications 誌に掲載されました。 →発生医学研究所ホームページ「New Press」での紹介記事
マウスの腎臓前駆細胞の起源が通説の初期中間中胚葉ではなく、下半身 の元となる体軸幹細胞様の細胞であることを見出し、実際にマウス胎児から採取したこの細胞から、腎臓前駆細胞を作るのに必要な5種類 の成長因子を特定しました。さらにこの5因子を適切な組み合わせと濃度で5段階に分けて投与することで、マウス ES (embryonic stem : 胚性幹) 細胞 およびヒト iPS (induced pluripotent stem: 人工多能性幹) 細胞の両方から腎臓前駆細胞を試験管内で作製することができました。そしてこれらの腎臓前駆細胞をさらに培養することで、糸球体 と尿細管の両方を伴った3次元の腎臓組織を試験管内で再構築しました。本研究は、腎臓の元になる細胞が胎内で形成される仕組みを明らかにするとともに、世界で初めて試験管内での3次元腎臓組織の構築を実現したものです。こ の腎臓組織が機能するためには、さらなる細胞の成熟化と尿を産生・排出する仕組みが必要ですが、本研究はこれまで固く閉ざされていた腎臓の再生医療の扉を 開く 大きな一歩と言えます 。またこの方法を元に、腎臓の病気を試験管内で再現できる可能性があり、病因の解明と創薬開発につながることが期待されます。本研究成果は、 2013年12月13日Cell Stem Cell 誌に先行掲載されました。




