




マウス胚性幹細胞(embryonic stem cells: ES細胞)は、着床前の胚盤胞の内部細胞塊に由来する、多能性幹細胞である(Evans and Kaufman, Nature, 1981; Martin, Nature, 1981)。マウスES細胞は、サイトカインLIF(leukemia inhibitory factor)依存的に、多能性を維持したまま細胞分裂を繰り返す。このような多能性幹細胞の自己複製には、多くの遺伝子機能が関与している。とりわけ、多能性幹細胞に高い特異性をもって発現する転写因子は、その中心的役割を果たしている。Oct3/4は、1990年に胚性癌細胞(embryonal carcinoma cells: EC細胞)で未分化状態特異的に発現する転写因子として同定された (Okamoto et al, Cell, 1990; Scholer et al, Nature, 1990; Rosner et al, Nature, 1990)。我々は、その機能がマウスES細胞の自己複製に必須で、その機能廃絶は栄養外胚葉(trophectoderm)への分化を誘導することを見出した(Niwa et al, Nat Genet, 2000)。さらに、Oct3/4と協調して機能する転写因子Sox2も、その機能廃絶は栄養外胚葉への分化を誘導することを報告した(Masui et al, Nat Cell Biol, 2007)。しかし、単一の転写因子の機能廃絶が、このような劇的な細胞分化を齎すことは寧ろ稀であり、高い未分化状態特異的発現を示すにも拘らず、その機能廃絶がなんら表現型を示さないこともしばしば認められる(例えばRex1/Zfp42やZscan10: Masui et al, BMC Dev Biol, 2008; Yamane et al, BBRC, 2015)。

では、単独の遺伝子破壊ではなんら異常を示さないような転写因子は、マウスES細胞の多能性維持に全く寄与していないのだろうか?マウスES細胞は、LIF依存性に自己複製するが、転写因子の人為的発現により、LIF要求性を解除できる。Nanogはこのような性質を示す転写因子として同定された(Mitsui et al, Cell, 2003; Chambers et al, Cell, 2003)。その後、我々は、同様の機能を持つ転写因子として、Klf4とTbx3を同定し、これらの転写因子がLIFシグナル入力を遺伝子発現へと変換する並列経路を構成していることを明らかにした(Niwa et al, Nature, 2009)。このような並列経路の存在は、一つの転写因子機能の欠損を代償する機構として機能しうる。さらに、Oct3/4標的遺伝子の網羅的解析から、Klf4, Tbx3, Nanog、そしてSox2やOct3/4自身も、Oct3/4によってその発現を正に制御されていることがわかった(Sharov et al, BMC Genomics, 2008など)。従って、これらの転写因子群は、シグナルを伝達する単方向性のカスケードではなく、双方向性に制御関係を持つネットワークを構成し、機能していると考えられる。
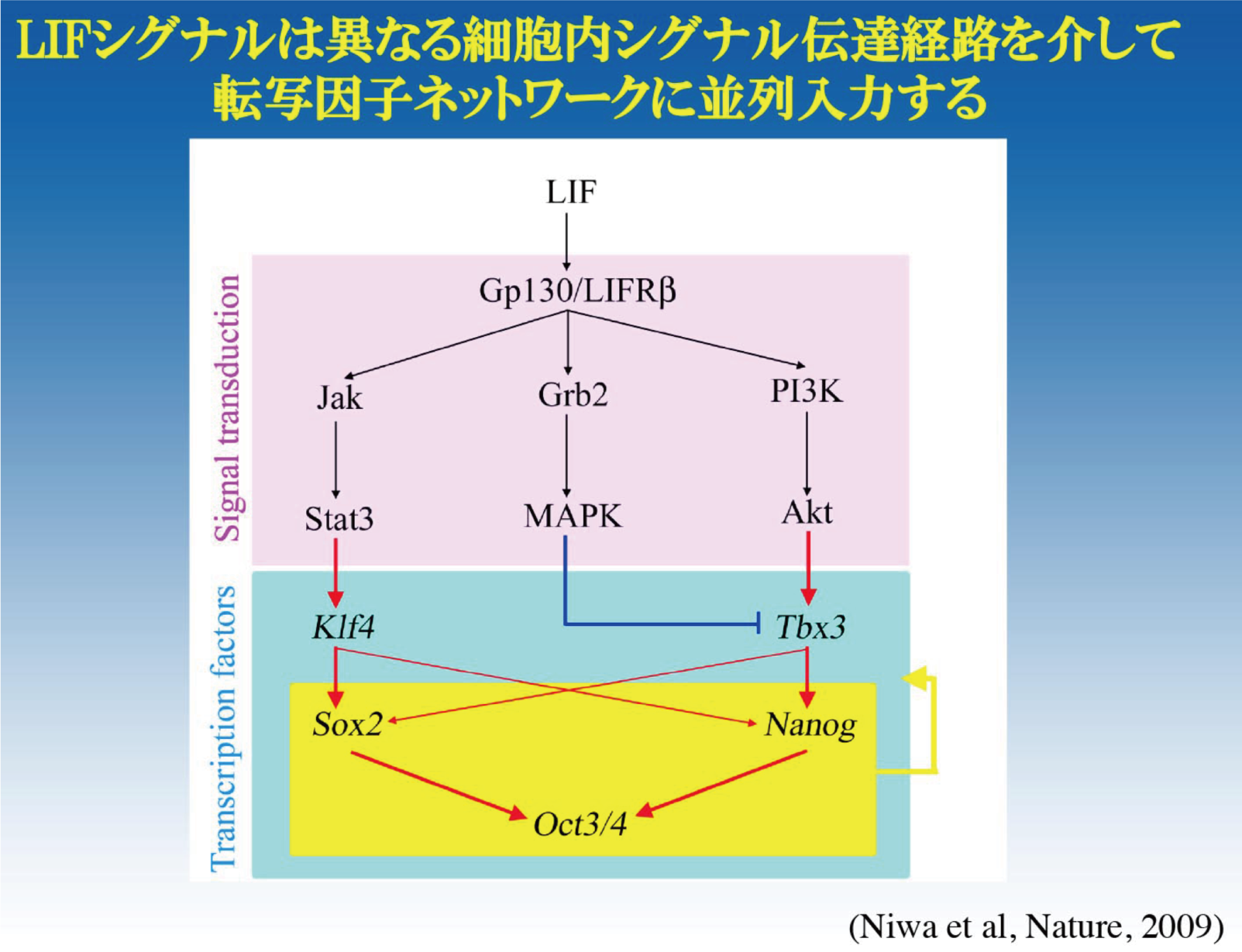
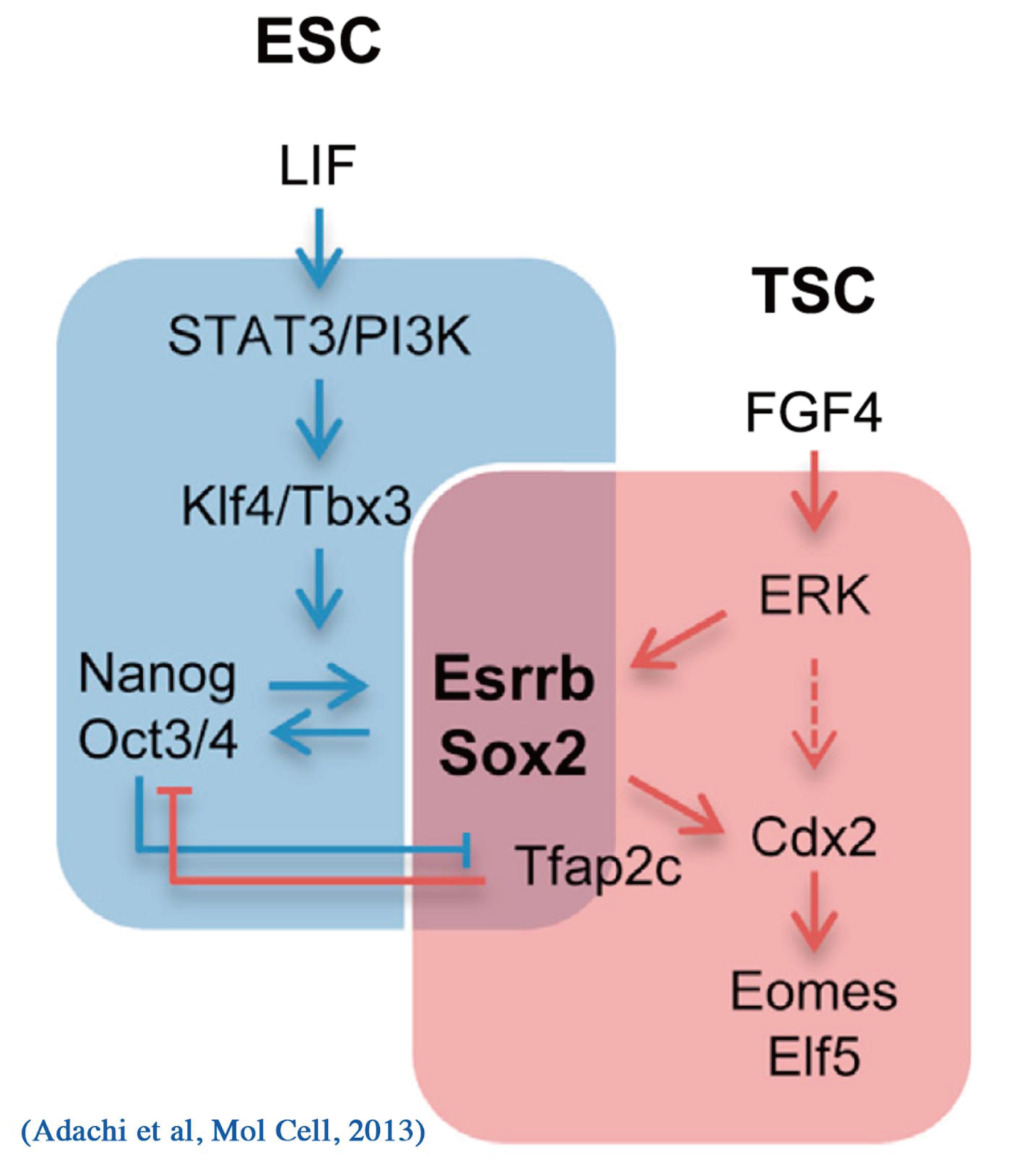
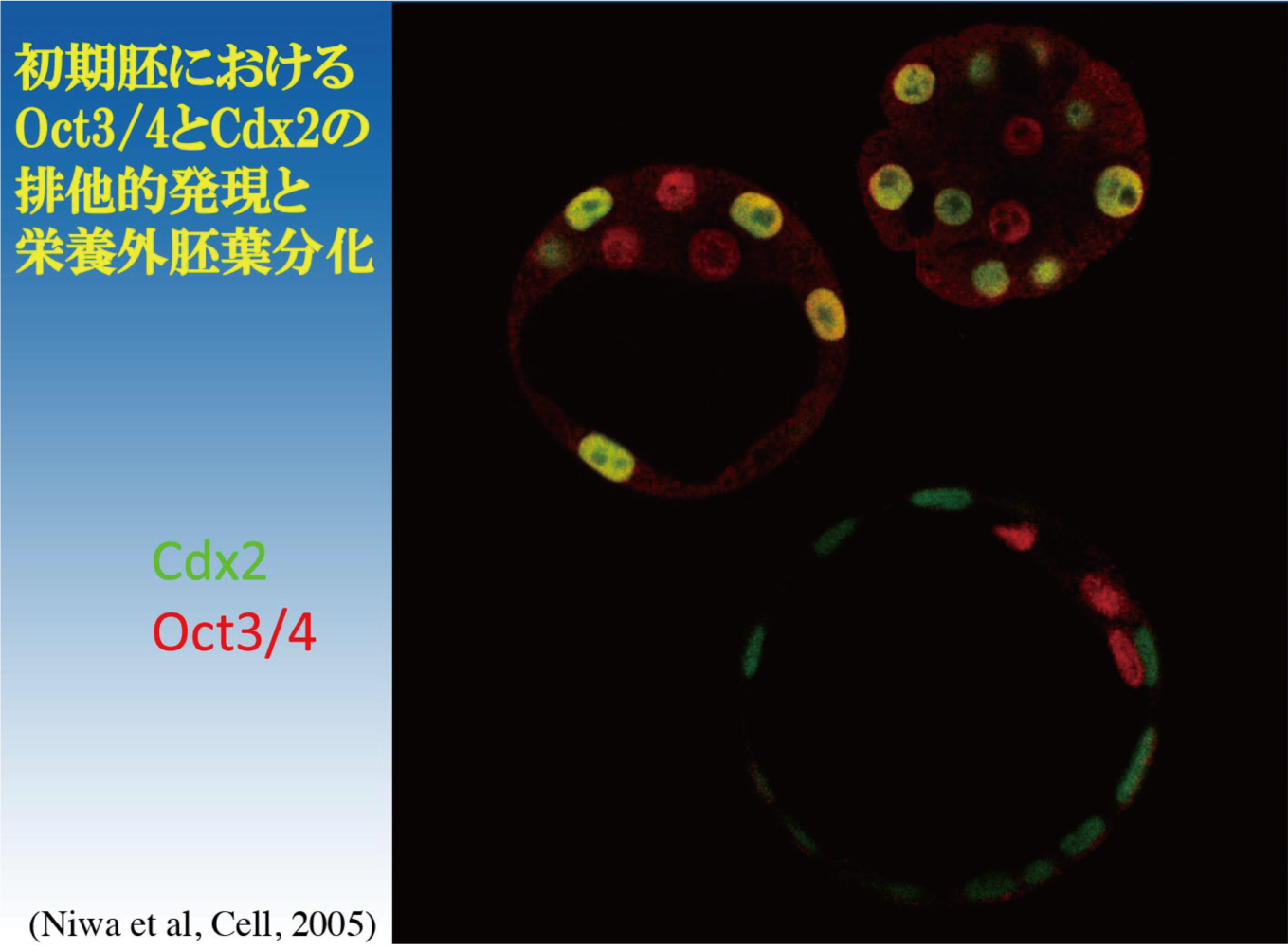
細胞種特異的に発現する転写因子群がネットワークを構成し、その全体の機能が細胞の分化表現型を規定しているとするネットワークモデルは、ゲノムワイドな解析からも支持されつつある。このモデルに従うならば、細胞の分化に伴い、転写因子ネットワークの遷移が起こることになる。マウスES細胞は、Oct3/4の機能破壊により栄養外胚葉へと分化するが、FGF4を含む培地でマウス線維芽細胞のフィーダー上で誘導すると、栄養外胚葉幹細胞(trophoblast stem cells: TS細胞)になる(Niwa et al, Nat Genet, 2000)。このとき発現が誘導される転写因子の機能検定から、Cdx2単独の人為的発現が同様の分化を誘導するのに十分であることがわかった(Niwa et al, Cell, 2005)。Oct3/4とCdx2は相互抑制機構を有することにより、その排他的発現が担保され、多能性細胞と栄養外胚葉の分化運命決定に寄与すると考えられる(Niwa et al, Cell, 2005; Niwa, Development, 2007)。これらの転写因子の発現操作によるES細胞からTS細胞への分化過程において、Sox2の発現は、ほぼ一定に維持される。では、その機能はどうなっているのだろうか?我々はクロマチン免疫沈降法により、分化過程におけるSox2のゲノムへの結合状態の変化を解析した。その結果、Sox2の結合は分化に伴い大きく変化すること、そしてその変化には、Sox2と協調する転写因子が分化に伴いOct3/4からTfap2cへと置き換わることが寄与していることを見出した(Adachi et al, Mol Cell, 2013)。この発見は、転写因子ネットワークの遷移が、それらを構成する個々の転写因子の機能を決定することを意味するものである。
細胞分化運命の決定に重要な役割を果たすと考えられる機構として、転写因子ネットワークのようなジェネティックな遺伝子発現制御機構とともに、ヒストン修飾やDNAメチル化によるエピジェネティックな制御機構がある。ES細胞では、ゲノムを担う染色体の構造は全般的に開いた状態にあり、そのことが分化に伴う全ての遺伝子の発現可能性を担保していると推測される(Niwa, Development, 2007; Niwa, Genes Dev, 2007)。しかし、この状態は細胞分化に伴い破綻し、選択された分化運命に必要でない遺伝子領域は閉じた状態(ヘテロクロマチン)となり、不活性化される。このような制御は、転写因子の結合標的配列の許容性(accessibility)を低下させることにより、転写因子の機能を限定し、いわゆるepigenetic landscapeとして分化の可逆性を抑制すると考えられるが、それを実験的に証明することは困難であった。我々は、転写因子Gata4の人為的活性化によるES細胞から原始内胚葉細胞への分化誘導系(Fujikura et al, Genes Dev, 2002; Shimosato et al, BMC Dev Biol, 2007)を用いて、新規DNAメチル化酵素Dnmt3a, Dnmt3bによるGata4標的配列近傍のメチル化が、分化細胞におけるGata4標的結合能を抑制することにより、その機能を制御していることを解明した(Oda et al, PLoS Genet, 2013)。これは、エピジェネティック制御が、転写因子ネットワークの機能を制御し、協調的に分化可逆性を規定することの証明に成功した、極めて重要な研究成果と考えている。
我々のこれまでの研究は、組織特異的転写因子は相互に制御し合うことにより転写因子ネットワークを構成して機能していること、そしてその活性は、液性因子などからの外部シグナル入力と、転写因子によって制御される標的遺伝子の結合許容性を規定するエピジェネティック制御機構との関係により規定されていることを明らかにした。しかし、これらの関係がどのようにして細胞分化を制御しているのかについては、まだまだ理解が及んでいない。例えば、特定の遺伝子領域に起こるエピジェネティック制御の標的特異性は、どのように規定されているのだろうか?特定のゲノム配列を認識できるのは転写因子であるからには、転写因子がエピジェネティック制御因子を特定の標的配列へと導いているはずなのだが、その機構は未だ明らかにはされていない。そして、変化したエピジェネティックな状態は、転写因子の結合を変化させるはずだ。このような相互作用のループが、細胞分化過程を規定し、その厳密性を保証していると考えられる。これはいわば、発生過程においてゲノム情報を読み出すための法則でもあるだろう。このような法則を司る原理を、ES細胞を用いた研究から引き出すこと、これが我々の研究の究極の目標である。




