




Yasufumi Yakushiji, Shingo Nishikori, Kunitoshi Yamanaka, and Teru Ogura (2006) Mutational analysis of the functional motifs in the ATPase domain of Caenorhabditis elegans fidgetin homologue FIGL-1: Firm evidence for an intersubunit catalysis mechanism of ATP hydrolysis by AAA ATPases. J. Struct. Biol. 156, 93-100.
AAA ATPase は、一般にリング状の 6 量体を形成して機能する。 ATP はサブユニットの境界領域に結合し、その加水分解には 3 つの機能的モチーフ( Walker A, Walker B および SRH )が関与することが提唱されている。細胞複製分野(小椋 光教授)の薬師寺 恭文(元博士課程大学院生)・錦織 伸吾(学振特別研究員)らは、線虫 FIGL-1 タンパク質の ATP 加水分解機構の詳細な解析を行った。まず野生型 FIGL-1 タンパク質と、 Walker A 変異体( K362A )、 Walker B 変異体( E416A )、および SRH 変異体( N461A と R471A )を調製した。予想されたように、いずれの変異体でも ATPase 活性は検出されなかった。ところが 2 種類の変異体を混合すると、組み合わせによっては ATPase 活性が検出された。例えば E416A と N461A の組み合わせ(図 A )では ATPase 活性は検出されなかったが、 E416A と R471A の組み合わせ(図 B )では活性が検出された。活性が検出された組み合わせでは、いずれかのサブユニット間に 3 つのモチーフのすべてが揃う。この結果は、提唱されていたサブユニット間触媒機構を強く支持している。おもしろいことに、 K362A と R471A の組み合わせ(図 C )では、あるサブユニット間に 3 つのモチーフが揃うにもかかわらず、 ATPase 活性が検出されない。この結果から、 3 つのモチーフが揃うことに加え、隣りの ATP 結合部位への ATP 結合によるサブユニットの構造変換が必要であるという、あらたなサブユニット間協調機構を提唱した。これらの研究成果は、 J. Struct. Biol. 誌 10 月号に発表された。
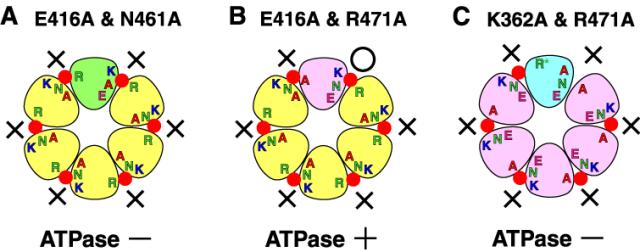
図 2 種類の変異体混合による ATPase 活性回復の説明図
ATP を赤丸で示す。○は ATP 加水分解できたこと、×はできなかったことを示す。 (A) E416A (黄色)と N461A (緑色)の組み合わせでは、どのサブユニット間にも 3 つのモチーフが揃わず ATP 加水分解は起こらない。 (B) E416A (黄色)と R471A (桃色)の組み合わせでは、あるサブユニット間に 3 つのモチーフが揃うことにより、 ATPase 活性があらわれる。 (C) K362A (青色)と R471A (桃色)の組み合わせでは、あるサブユニット間に 3 つのモチーフが揃うにもかかわらず、隣りの位置に ATP が結合していないため R471 が機能的でなく、 ATP 加水分解が起こらない。
