









当研究室では,AAAファミリータンパク質の分子・細胞・発生生物学的研究を行っています。大学院生[修士・博士]も広く募集しております。お問い合わせは山中准教授<yamanaka(at)kumamoto-u.ac.jp>まで。
(1) 線虫のAAAタンパク質の細胞機能と発生における役割の解析およびこれらの変異に起因する疾患モデルの作製と病態解析。
(2) タンパク質凝集体病の線虫疾患モデルの開発と治療候補薬の探索

AAAファミリータンパク質の研究
細胞複製分野は、 AAAファミリータンパク質(ATPases associated with diverse cellular activities)の共通分子基盤と多彩な細胞機能の解明をめざしています(図1)。AAAファミリータンパク質は、リング状6量体を形成して機能するATP依存性の分子シャペロンです(図2)。タンパク質をアンフォールド(unfold)したり、タンパク質複合体を脱会合(disassembly)したり、タンパク質凝集体を脱凝集(disaggregate)する活性をもつのが特徴です。真核生物は20数個のほぼ同じセットのAAAタンパク質をもっています。近年、AAAタンパク質に起因する、あるいは関連するヒト遺伝性疾患(ペルオキシソーム病、遺伝性痙性対麻痺、ポリグルタミン病、骨パジェット病と前側頭葉型痴呆を伴う家族性封入体筋炎など)も相次いで報告され、医学・薬学的見地からも注目されています(表1)。AAAタンパク質の研究は、きわめて重要な未解決の問題が残されているチャレンジングでホットな分野です。AAAファミリータンパク質についてもっと詳しく知りたい方は、 AAAファミリータンパク質のホームページ を参照してください。
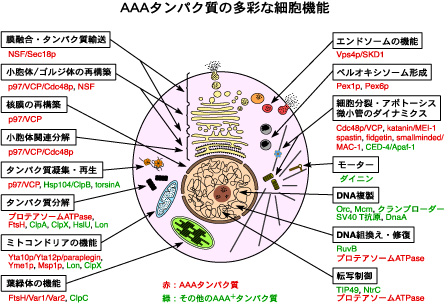
図1.AAAタンパク質の多彩な細胞機能
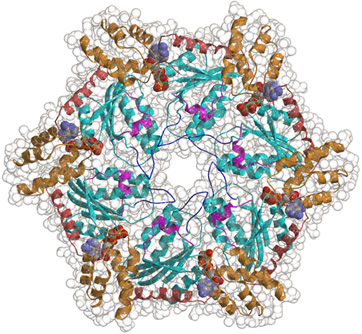
図2.AAAタンパク質のリング状6量体構

表1. AAAタンパク質に起因する疾患
AAAファミリータンパク質の構造と分子機構の解明
AAAタンパク質の基本構造や共通分子機構を解明することは、この新しいタイプのシャペロンをよく理解するための第一歩であり、AAAタンパク質の多彩な細胞機能を解明するためにも重要です。大腸菌のAAAプロテアーゼFtsHや線虫のAAAタンパク質などを使って、生化学的手法や分光学的手法により反応機構や構造変化を解析しています。
線虫の AAAタンパク質の細胞機能の解析とヒト疾患モデル
線虫(図 3)は、多細胞動物としての基本的体制を備えた最小の生物の一つで、実験動物としての様々な利点があります。培養が簡単で世代時間が短いこと、遺伝的手法が確立していること、細胞が透明で観察しやすいこと、959個の体細胞からなり、それらの細胞系譜が明らかにされていること、神経細胞は302個あり神経回路網が明らかにされていることなどです。線虫はヒトとほぼ同じセットの24個のAAAタンパク質をもち(図4)、同定されているヒト遺伝性疾患に関わるすべてのAAAタンパク質のホモログが存在します。これらの遺伝性疾患に関連したAAAタンパク質に注目して線虫のAAAタンパク質の細胞機能を解析しています。
図3.線虫
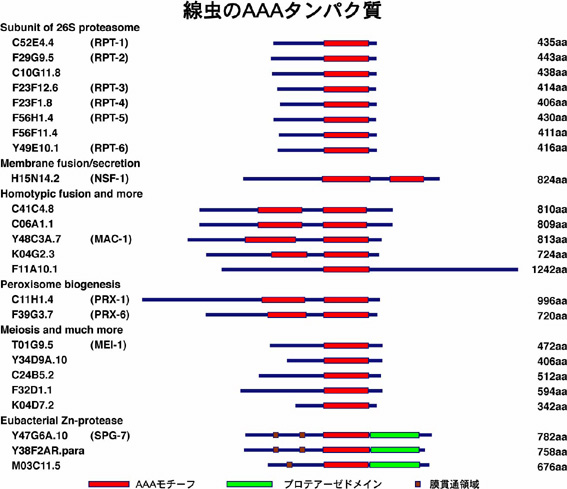
図4.線虫のAAAタンパク質
現在、研究対象としている AAAタンパク質と研究テーマは以下の通りです。
AAAプロテアーゼFtsHの解析:
FtsHは真正細菌に普遍的に存在する膜結合型のATP依存性プロテアーゼです(図5)。大腸菌のFtsHの細胞機能についての解析がほぼ終わり、現在は基質タンパク質のunfolding、translocation、分解の分子機構を解明する研究を行っています。
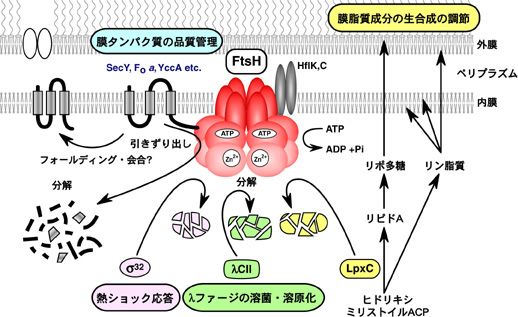
図5. AAAプロテアーゼFtsH
Parapleginの解析:
真核細胞のミトコンドリアの内膜には FtsHホモログが複数個存在します。活性部位がマトリクス側にあるm-AAAプロテアーゼと膜間スペース側にあるi-AAAプロテアーゼがあり、膜タンパク質の品質管理を行っています(図6)。線虫には、2個のm-AAAプロテアーゼと1個のi-AAAプロテアーゼがあります。ヒトm-AAAプロテアーゼの変異は遺伝性痙性対麻痺(hereditary spastic paraplegia)の原因となり、parapleginとよばれます。遺伝性痙性対麻痺は特に下肢の進行性の麻痺をおもな症状とする神経変性疾患です。 線虫のparapleginホモログの細胞機能、発生における役割、運動能などについて解析するとともに、精製タンパク質を用いてFtsHと比較しながら解析しています。
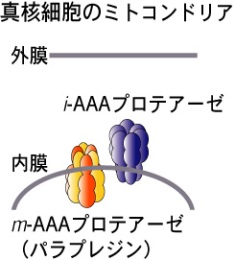
図6. ミトコンドリアのAAAプロテアーゼ
Spastinの解析:
遺伝性痙性対麻痺の原因因子として、 parapleginに加え、もう一つのAAAタンパク質spastinが同定されています。このspastinはミトコンドリアには局在しないし、プロテアーゼでもありません。微小管のダイナミクスに関わることが示唆されています。線虫のspastinホモログの細胞機能と微小管への作用を解析しています。
Fidgetinの解析:
Fidgetinは、マウスの頭部(特に眼と耳)の形成が異常となるfidget変異の原因因子として同定されました。Fidgetinは構造的にspastinに極めて類縁のAAAタンパク質です。線虫のfidgetinホモログの細胞機能と精製タンパク質のin vitro解析を行っています。
p97/VCPの解析:
p97/VCPは、細胞分裂後の小胞体、ゴルジ体、核膜の再構築に働く因子として同定されましたが、その他にも様々な細胞機能に働くことが示唆されています。小胞体関連分解(ERAD)への関与やポリグルタミン結合能などです。骨パジェット病と前側頭葉型痴呆を伴う家族性封入体筋炎の原因因子としても同定されています。ポリグルタミン鎖を含むタンパク質は凝集体を形成しやすく、神経変性を引き起こすポリグルタミン病の原因になります。線虫には、p97/VCPホモログが2個存在し、それぞれの機能分担を解析するとともに、ポリグルタミン病モデル線虫を作製し、ポリグルタミン凝集体への作用を解析しています。また、p97/VCPの多機能性に関わるアダプタータンパク質の探索を行っています。さらに、近縁のAAAタンパク質についても解析しています。精製タンパク質によるポリグルタミン凝集体の脱凝集(disaggregation)についてもin vitroで解析しています。
Kataninの解析:
Kataninは微小管上で会合し、微小管を切断する(会合しているチューブリンをdisassembleする)活性があります。ヒトkataninの微小管への作用をin vitroで解析しています。
結晶構造は York大 Wilkinsonグループとの共同研究で、1分子イメージングは学習院大 西坂グループとの共同研究で行っています。AAAタンパク質およびその基質の動的変化を高速原子間力顕微鏡で観察する共同研究を金沢大 安藤グループと進めています。合成ポリペプチドを用いた解析には鳥取大 櫻井グループの協力を得ています。
以上の研究は、科研費【特定領域研究「タンパク質の一生」(領域代表 吉田賢右);基盤研究】、科学技術振興機構 CREST【「タンパク質のナノダイナミクス高速撮影装置の開発」(代表者 安藤敏夫)】などの研究費により行われています。








