



1-1. FtsHプロテアーゼの構造と分子機構
大腸菌の増殖に必須の ftsH遺伝子を同定し、FtsHの構造と機能について研究してきた。FtsHはN末端側で膜を2回貫通する膜内在性タンパク質で、細胞質側に ATPaseドメインとプロテアーゼドメインをもち(Tomoyasu et al., 1993; 1995)、リング状オリゴマーを形成する(Shotland et al., 1997; Akiyama et al., 1998)。このオリゴマー形成には膜貫通部位と膜外ドメインが重要である。AAA ATPaseはそのATPaseドメイン内にこのファミリーを特徴づける領域(SRH: second region of homology)を持つが、その機能について解析し、この領域がATPase活性に重要であることを明らかにし、AAA ATPaseによるATP加水分解機構について、隣り合う2つのサブユニットが協同してATPを加水分解するという「分子間触媒」モデルを提唱した(図1)(Karata et al., 1999; 2001)。FtsH ATPaseドメインの結晶化に成功し、その結晶構造を解いた(Krzywda et al., 2002)。これにより、「分子間触媒」モデルの構造的基盤が得られた。FtsHの 6量体リング構造の中央の孔に位置する保存された芳香族残基が基質ポリペプチドのtranslocationに重要であることを証明した(図2)(Yamada-Inagawa et al., 2003)。これは、 基質タンパク質をATP依存的にunfoldし、 リングの孔を通過させるという「糸通し」モデルを支持する。種々の基質について、その分解の方向性やエネルギーコストを詳細に解析した(Okuno et al., 2004)。
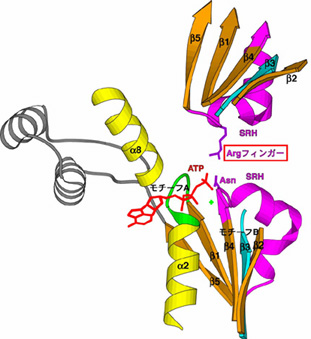
図1. AAA ATPaseによるATP加水分解の「分子間触媒」モデル
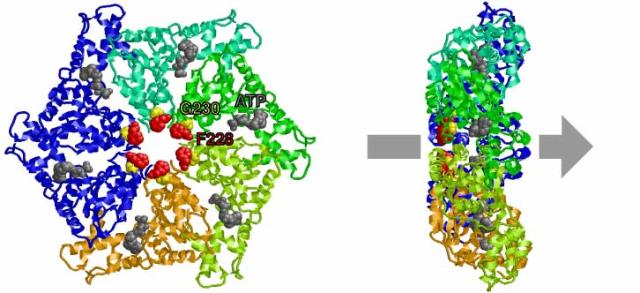
図2. AAA ATPaseリングの孔の入口で保存された芳香族残基
1-2. FtsHプロテアーゼと分子シャペロンによる熱ショック応答制御
大腸菌の熱ショック応答は熱ショック転写因子σ 32の量と活性の調節がその制御機構の中心である。FtsHがσ32を特異的に分解するプロテアーゼであることを発見した(図3)(Tomoyasu et al., 1995)。一方、分子シャペロンDnaKシステムは、σ32の分解と不活性化の両方に関与する(Tatsuta et al., 1998; Tomoyasu et al., 1998)。
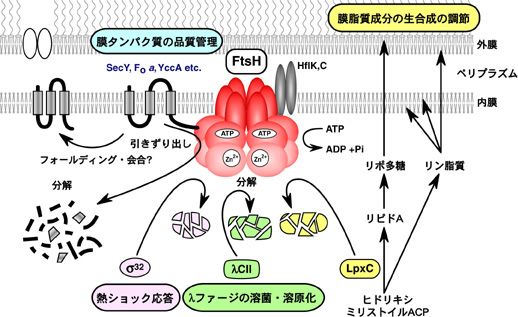
図3. FtsHの細胞機能
1-3. 膜の形成と品質管理におけるFtsHの機能
膜の主要構成成分であるリン脂質とリポ多糖は共通の前駆物質からアシル基を受け取るので、その合成は競合的である。 FtsHはリポ多糖の合成経路の酵素LpxCを分解し、その量を調節してリポ多糖とリン脂質の合成バランスを制御することを明らかにし、これがFtsHプロテアーゼの細胞増殖に必須の機能であることを突き止めた(図3)(Ogura et al., 1999)。FtsHは、膜タンパク質複合体の品質管理にも関与することが報告されており、これらを併せ考えると、FtsHは膜形成・膜動態を総合的に制御する調節因子である。ftsH変異株では、ミニFプラスミドの分配異常が起こる(Inagawa et al., 2001)。
2-1. ミトコンドリアに局在するFtsHホモログの解析
線虫の2つの m-AAA プロテアーゼのうち、Y47G6A.10の阻害では成長の遅 延や運動能に異常がみられ、組織化学的アッセイや電顕観察(神戸大 洲崎グループの協力)によりミトコンドリアの異常を認めた。線虫の m-AAA プロテアーゼを精製し、そのATPase活性、プロテアーゼ活性を解析している。
2-2. Fidgetinホモログの解析
線虫の fidgetinホモログの欠損は生殖腺形成不全・不稔になることを観察している。Fidgetinホモログを精製し、in vitroでATPase活性を解析し、ATPase活性に重要なシステイン残基を同定した(Yakushiji et al., 2004)。また、部位特異的変異体を用いた解析から、ATP加水分解の「分子間触媒」モデルを支持する結果を得たほか、協調性に関する新たな知見も得た。
2-3. p97/VCPホモログの解析
線虫に存在する2個の p97/VCPホモログを同時に阻害すると胚致死になる。致死卵の電顕観察により、細胞内に多数の空胞様構造を認めた。これは膜融合や小胞体機能が阻害されたことによると考えられる (Yamanaka et al., 2004) 。線虫で種々の長さのポリグルタミン鎖を発現すると、鎖長が40を越えると凝集体が形成され、ヒトポリグルタミン病のモデルとなる(図4)。このとき p97/VCPホモログを共発現するとポリグルタミン凝集体が部分的に減少することを観察した(Yamanaka et al., 2004)。現在、凝集体抑制機構の詳細をin vivo, in vitroで精力的に解析している。また、 p97/VCPホモログの一つC06A1.1の欠失変異体では、受精卵の数が減少し、この現象が精子形成から卵母細胞形成へのスイッチング異常によることを見出した。
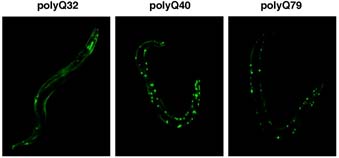
図 4.線虫のポリグルタミン病モデル
2-4. Spastinホモログの解析
Spastinは微小管と相互作用し、katanin同様、微小管をする活性を持つことが最近報告された。線虫のspastinホモログもチューブリンに結合することを見出した。Spastinホモログを欠失した線虫は、卵形成が異常になるなどの表現型を示す。




