



石黒研究室(染色体制御分野)は、千葉大学医学研究院・疾患生命医学へ転出致しました。
Twitter https://twitter.com/IshiguroL
着任の挨拶
当分野は2016年9月に立ち上がった研究室です。私は長年、任期付きのポスドク、助教、講師を経験して参りました。そして今もなお、タイミング良く職を得て研究を継続していくことの難しさを強く認識しております。発生研において染色体制御分野を新設する機会を得たことはこの上ない僥倖であったと思います。まさにドイツの社会学者マックス・ウェーバーの著書「職業としての学問」(訳:岩波文庫)に出てくる一節が思い起こされます。「若い学者にとって職業としての学問は就職および昇進において「僥倖」に支配されている。・・・・・」そこでは、当時の大学制度において、職を得ることは思いもかけぬ偶然によって支えられていること、また同僚の就職や昇進をよそ目に己の精神を如何にコントロールし学問に没頭できるかが、職業としての学問を続ける上での資質であると説いております。今から100年も前に記されたものですが、いまの自分を取り巻く境遇と相重なるものを感じております。
近年、大学院進学率は軒並み減少傾向にある状況です。私に微力ながら出来ることがあるとしたら、まず研究の場に触れるきっかけを作ること、そして考え思い巡らす研究活動が収入や社会ステータスのような尺度では計り知れないものであることを伝えることだと思います。震災に見舞われながらも、底力を以て起ち上がった発生研から大いに発信できるような研究と教育活動に邁進する所存です。
当分野への研究参加
研究室は小規模ですが、濃密かつ出口のはっきりした実効性のある研究・教育活動を目指しています。我々は分子生物学、細胞生物学、生化学、発生生物学に幅広く跨がる領域を指向しており、出身学部やこれまでの研究バックグラウンドを問いません。むしろ無知であることを恥ずかしがらないこと、経験にとらわれず柔軟であること、怖いもの知らずであることは、新しいことに挑戦する研究者としての重要な資質だと思います。研究は頭を使うことも大事ですが、時にひたむきに気合いと根性で突き進む力業も大事であると考えます。意欲ある大学院生、研究員、基礎研究に関心のある臨床医、社会人経験者などの方々の積極的な参加を歓迎致します。大学院生は、大学院医学教育部 医科学専攻(修士課程)または医学専攻(博士課程)から受け入れております。日本学術振興会特別研究員としての参加についてもご相談に応じます。当分野の研究内容に興味のある方はお問い合わせ下さい。
研究目標
当染色体制御分野では高等動物の減数分裂における染色体構築とその制御のメカニズムについて研究を推進します。染色体構成の次世代への正確な継承と初期胚の正常発生の観点から、減数分裂は生殖細胞に特有かつ極めて重要なイベントです。とりわけ、当分野では以下に記す3つの角度から基礎研究を行います。内容的には高齢出産、少子化の観点で、医学分野のみならず社会的にも強くアピールできる研究課題であると考えます。
いずれの研究テーマに関しても、新規の未解析因子を見つけ出すことを重視します。分野のパラダイムシフトには、missing linkを埋める新規因子の同定によって新しく現象を説明することが、重要な研究スタンスであると考えます。これまでの研究経験から、生殖細胞を見渡しただけでもデータベースに眠る未解析因子は相当数残されており、まだまだ発掘の余地があると考えられます。
生殖細胞の研究を行うにあたって念頭におくべきことがあります。それは、あらゆる生物種に共通する原理的な保存性と、生物種に固有のメカニズムが存在するということです。先行研究から、高等動物の減数分裂に特有で他の種には見られない現象があることや、アミノ酸配列上はまったく相同性のないものが高等動物とそれ以外の種で似た役割を果たす事例が見出されています。特に、生殖細胞には種にユニークな機構が備わっている例が多く知られることからも当分野では主にマウスを用いた研究を行います。高等動物では、酵母、ショウジョウバエ、線虫のような変異体取得の遺伝学的スクリーニング手法が容易に適用できないため、当分野の目指すハードルは高く、また国際的にも攻め倦んでいる状況です。しかしながらヒトの不妊・不育などのメカニズムを考えた場合には、ヒトの疾患の原因解明に道筋をつけるためにも敢えてマウス個体や胚性幹細胞を用いることに本研究を行う意義があると考えます。発生研という質の高い研究環境と近隣の優れた研究施設の地の利を活かして、究極的には医学系大学院・臨床講座との連携によって生殖医療にも資するように、減数分裂の分子機構の解明を目指します。
研究内容
(1)体細胞増殖から減数分裂への切り替え制御に関する研究
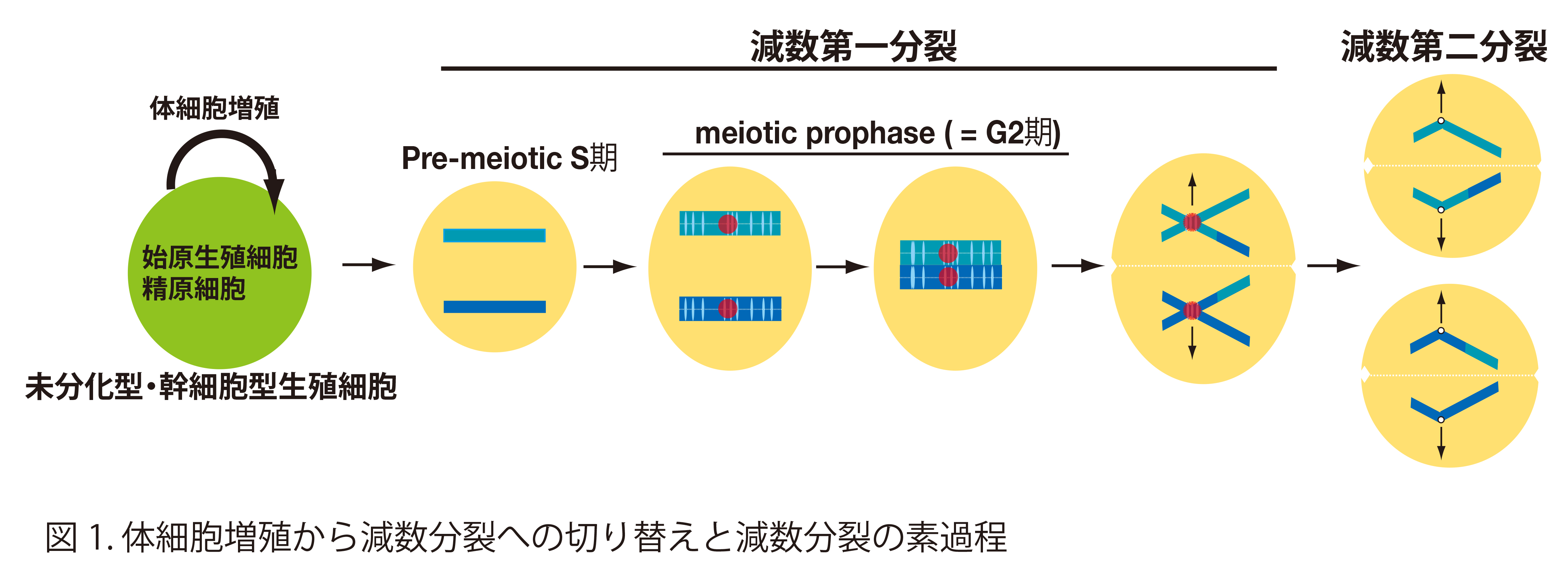
マウスの場合、未分化型・幹細胞型の生殖細胞は体細胞型の増殖を経た後に、その一部の集団がspontaneousに減数第一分裂のpre-meiotic S期へと進行する(図1)。この時、体細胞型増殖から減数分裂型cell cycleへの切り替えが起きていると推測されるが、その分子機構は多くの点が不明のままである。特にこの減数第一分裂への移行期においては染色体構造が減数分裂仕様に再構成されると推測され、染色体制御を理解する上でキーとなるステージであるにもかかわらず材料の量的・数的な制限のため研究は膠着している。最近、我々は減数分裂の開始に決定的な役割を担う新規の転写活性化因子MEIOSINを同定した(Dev. Cell 52, 2020)。これを欠損させると細胞周期の維持に関与する体細胞型Cyclinの異所的発現やM期様染色体構造など体細胞様の特徴を示す細胞の蓄積を伴って、体細胞分裂から減数分裂への切替えが見られなくなる。MEIOSINはSTRA8と複合体を形成して減数分裂関連遺伝子の転写開始点近傍に結合して転写活性化に働くことが明らかとなった。これによって直接制御される標的には多くのhypothetical gene (ゲノムデータベース上にIDのみが付与されている機能不明遺伝子)が含まれることが判明している。これらMEIOSINの転写制御下に置かれている未解析の遺伝子には、減数分裂の進行に必要とされる未知のものが含まれる可能性がある。そこで本研究では減数第一分裂の制御を支える新規遺伝子の網羅的解析を行って体細胞分裂との違いを本質的に決定付ける減数分裂制御のメカニズムの解明を目指す。
(2)減数分裂型細胞周期と染色体制御に関する研究
本研究では体細胞型と減数分裂型の細胞周期の制御機構の違いを見出すことを課題とする。生殖細胞は通常の体細胞と同様の細胞周期の機構を巧みに転用しながらも、染色体構造に減数分裂特異性が与えられるようにプログラムされている。減数分裂を細胞周期調節という観点から体細胞と比較すると、両者は様々な点で異なっている。例えば減数第一分裂前期(meiotic prophase I)と呼ばれる時期は、通常の体細胞の細胞周期のG2期に相当する(図1)。このmeiotic prophase Iは、そのタイムスパンが通常の細胞周期G2期と比べて際だって長いこと(精母細胞でおよそ7-8日にも及ぶ)が特徴で、染色体上の様々なイベントを達成させる猶予を与える期間であると考えられる。さらに、第一分裂M期が完了しても次のS期が開始されずに直ちに第二分裂M期へ進行する点も、通常の細胞周期と極めて異なる特徴の一つである。このように細胞周期の調節は減数分裂仕様に大幅に特殊化されているが、この分子機構はほとんど理解されていない。
さらに、卵子形成過程においては胎生期に始まる減数第一分裂前期が出生前にいったん停止して、ヒトでは数年〜数十年にもおよぶ長期の休眠状態にはいる。この停止状態の卵子はその後、徐々に限定された数だけ排卵にともない減数第一分裂を再開する。したがって卵子には減数第一分裂前期の進行を長期に渡って抑制するメカニズムが存在することが推定されるが、減数分裂における雌雄性差の本質的なことはあまりわかっていない。
体細胞で見られる細胞周期調節が染色体の動態に決定的な役割を果たしていることからも明白であるように、減数分裂における細胞周期調節の観点から染色体制御を理解することが重要となろう。本研究では、細胞周期の進行に重要とされるユビキチンリガーゼ複合体やシグナル伝達系のタンパク質の減数分裂の素過程における働きを酵素・基質・調節の観点から研究する。
(3) 減数分裂におけるコヒーシンの制御
コヒーシンはS期で複製された姉妹染色分体がばらばらとならないように、それらの接着に働くタンパク質複合体である。さらに体細胞においては、コヒーシン複合体はインスレータータンパク質と強調して遠方のエンハンサーとプロモーターとを手繰り寄せるようにして、遺伝子発現の調節に働くことも知られている。
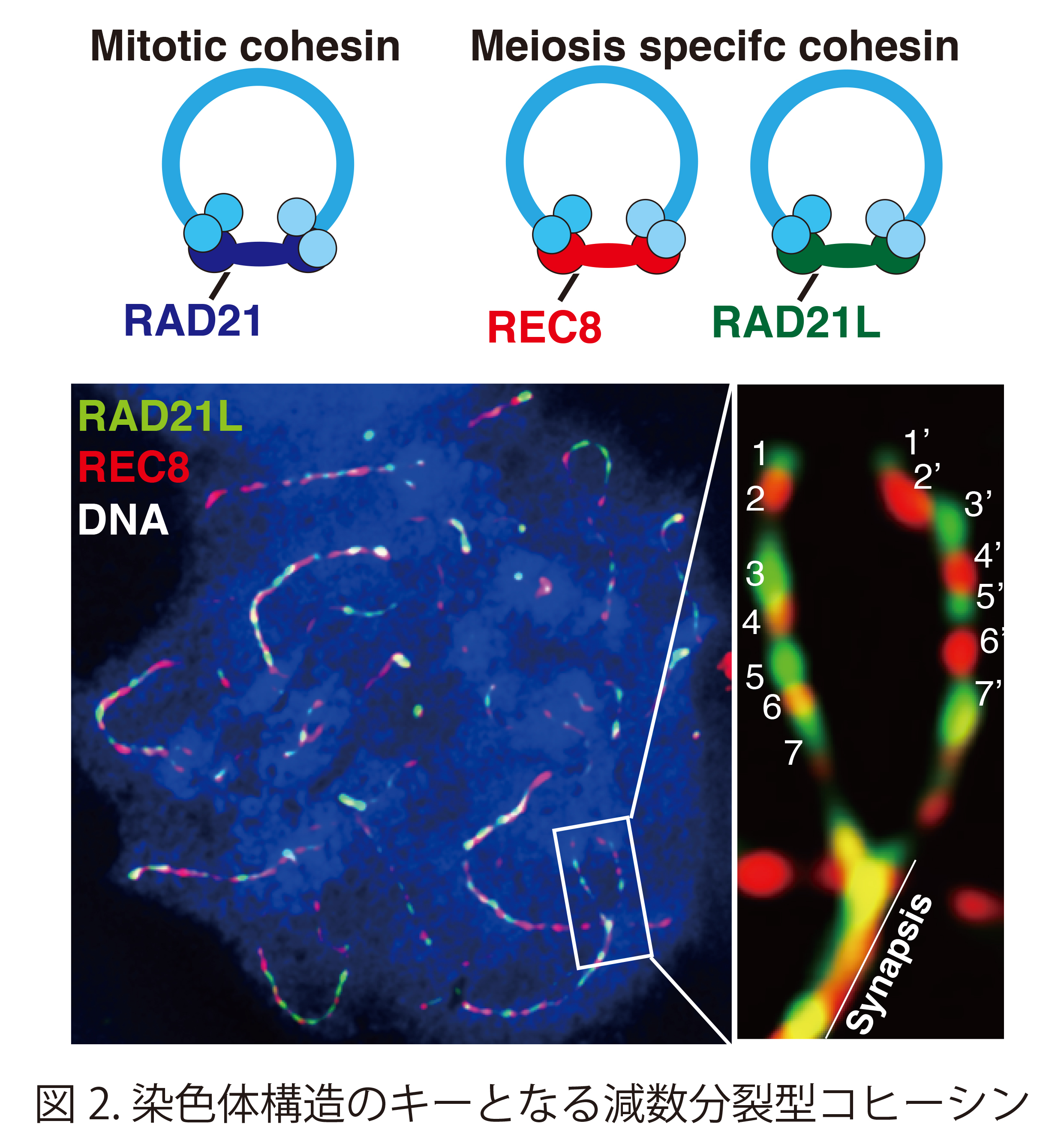
重要なことに、このコヒーシン複合体には体細胞型と減数分裂型の使い分けがある。すなわち、体細胞ではRAD21をサブユニットとして含むコヒーシン複合体があるのに対して、減数分裂では、RAD21LまたはREC8をサブユニットとして含むものが存在する。この減数分裂型コヒーシン複合体は、姉妹染色分体接着のみならず、体細胞では見られない減数分裂に特有の染色体構造の骨組みとしても極めて重要な役割を果たしている(図2)。さらに老化卵子において、コヒーシンの減少と加齢に伴う高頻度の染色体分配異常と相関があることも指摘されている。長期の休眠状態において卵子の染色体上のコヒーシンが安定に保持されるメカニズムが存在が推定されるが、その実体は明らかではない。
本研究課題では、この減数分裂型コヒーシンのローディングをライセンスする機構の解明を目指す。とりわけpre-meiotic S期は減数分裂型コヒーシンが染色体上に検出される最初の時期である。この時期には、染色体の減数分裂仕様に向けた再構成が行われているものと推測されるので、減数分裂型コヒーシンを制御する新たなクロマチン結合因子が見出されないか検討する。
当ラボでは以下の支援を受けて研究が行われています。
2025~2029年度 学術変革領域研究A(機動性ゲノム) 領域代表
2025年度 内藤記念科学振興財団
2024~2025年度 学術変革領域研究(生殖ライフスパン) 公募研究
2023年度 公益財団法人 上原記念生命科学財団
2023年度 アステラス病態代謝研究会ステップアップ研究助成
2023年度 三菱財団 自然科学研究助成
2023-2026年度 科研費基盤研究(A)
2022-2023年度 科研費挑戦的研究(萌芽)
2021-2024年度 AMED-PRIME 革新的先端研究開発支援事業「早期ライフ」
2021年度 武田科学振興財団 武田報彰医学系研究助成
2021年度 ノバルティス科学振興財団
2020-2022年度 科研費基盤研究(B)
2020-2021年度 科研費挑戦的研究(萌芽)
2020年度 第一三共生命科学研究振興財団
2020年度 内藤記念科学振興財団
2020年度 アステラス病態代謝研究会
2020年度 公益財団法人 上原記念生命科学財団
2020年度 住友財団 基礎科学研究助成
2019-2023年度 新学術領域計画研究(非ゲノム情報複製機構)
2019年度 新学術領域(配偶子インテグリティ)
2018-2019年度 科研費挑戦的研究(萌芽)
2017-2019年度 科研費基盤研究(B)
2016-2017年度 新学術領域研究(配偶子産生システムの制御)
2016-2017年度 科研費 新学術領域研究(生殖細胞エピゲノム)
2014-2016年度 科研費 基盤研究(C)
2017年度 武田科学振興財団 武田医学系研究助成 (継続)
2017年度 公益財団法人金原一郎記念医学医療振興財団
2016年度 公益財団法人山田科学振興財団
2016年度 上原記念生命科学財団
2015年度 武田科学振興財団
2015年度 持田記念医学薬学振興財団







