




Akinobu Onitake, Yuka Matsushita-Ishiodori, Ai Johjima, Masatoshi Esaki, Teru Ogura, and Kunitoshi Yamanaka , The C-terminal α-helix of SPAS-1, a Caenorhabditis elegans spastin homologue, is crucial for microtubule severing. Journal of Structural Biology, in press
緩徐進行性の下肢の痙縮と筋力低下を呈する神経変性疾患であるヒト遺伝性痙性対麻痺の原因因子は多数報告されているが、 spastin の変異もその一つである。 Spastin は、 ATP 依存的に微小管を切断する活性をもつ AAA タンパク質である。これまでに、分子細胞制御分野(小椋 光教授)では、 spastin の線虫ホモログ SPAS-1 も微小管切断活性を有することを報告している( Matsushita-Ishiodori et al., 2009 )。最近ショウジョウバエの spastin に関して、 C 末端領域の α- ヘリックスが ATPase 活性と微小管の切断に重要であることが報告された。
今回、 同分野の鬼武彰宣(博士課程大学院生)らは、線虫 SPAS-1 の C 末端領域の α- ヘリックス の重要性について詳細な解析を行った。野生型 SPAS-1 を培養細胞で発現させると微小管の消失が観察されるが、 C 末端 α- ヘリックス を欠失させた変異 SPAS-1 では微小管の消失が観察されなかった。また、 C 末端領域にプロリン残基を導入し、 α- ヘリックス を破壊した変異 SPAS-1 を発現させた場合も微小管の消失が観察されなかった(図)。次に C 末端 α- ヘリックス 欠失 SPAS-1 は ATPase 活性を示さないことを明らかにした。これまでに、 SPAS-1 の微小管への結合は、 SPAS-1 の安定な 六量体 形成を可能にし、その ATPase 活性を促進することが示唆されている。そこでゲル濾過クロマトグラフィーにより六量体形成能を調べたところ、 C 末端 α- ヘリックス 欠失 SPAS-1 も野生型 SPAS-1 と同様に安定な六量体を形成していることを確認した。次に構造変化に伴う内在性トリプトファン蛍光変化により ATP 結合能を解析したところ、野生型 SPAS-1 は ATP 結合能を有するのに対して、 C 末端 α- ヘリックス 欠失 SPAS-1 は ATP 結合能を失っていることを明らかにした。したがって、線虫 SPAS-1 の C 末端領域の α- ヘリックス は ATP 結合活性と微小管切断活性に重要な役割を果たしていることが示された。遺伝性痙性対麻痺の患者から見つかっている spastin の変異の 70 %は ATPase ドメインに集中しているが、 4 つの変異が C 末端 α- ヘリックス領域に 見つかっていることから、これらの患者では、 spastin の ATP 結合活性と微小管切断活性が欠損していることが推測される。これらの研究成果は、 J. Struct. Biol. 誌電子版に先行掲載された。
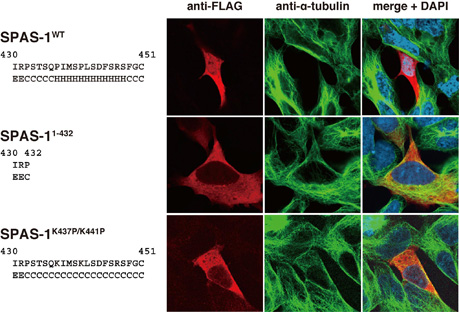
図:微小管切断における SPAS-1 の C 末端 α- ヘリックス の役割
野生型 SPAS-1 を HEK293 細胞で発現させると微小管の消失が観察された。 C 末端 α- ヘリックス 欠失 SPAS-1 及び K437/K441 にプロリン残基を導入した SPAS-1 では微小管の消失が観察されなかった。 FLAG タグを付加した SPAS-1 を抗 FLAG 抗体で、染色体を蛍光試薬 DAPI で、微小管を抗 α-tubulin 抗体でそれぞれ染色した。
