




Yuka Matsushita-Ishiodori, Kunitoshi Yamanaka, Hiroshi Hashimoto, Masatoshi Esaki, and Teru Ogura (2009) Conserved aromatic and basic amino acid residues in the pore region ofCaenorhabditis elegans spastin play critical roles in microtubule-severing. Genes CellsPublished Online: July 13 2009 .
遺伝性痙性対麻痺の原因因子の一つである spastin は, ATP 依存的に微小管を脱会合 (切断)することが報告されている。これまでに,分子細胞制御分野(小椋 光教授)の石躍(松下)由佳(博士課程大学院生)らは,線虫ホモログ SPAS-1 も微小管切断活性を有することを報告しており( Matsushita-Ishiodori et al . (2007)Biochem. Biophys. Res. Commun . 359 , 157-162 ),今回, SPAS-1 による微小管切断の分子メカニズムについて詳細な解析を行った。その結果, SPAS-1 の ATPase 活性は微小管により促進されることを見出し, SPAS-1 の N 末端領域 ( microtubule binding domain: MTBD )で tubulin と直接相互作用していることを明らかにした。また,ゲル濾過クロマトグラフィーにより SPAS-1 の 6 量体形成は濃度依存的に起こり, ATP を必要としないことを明らかにした。これらの結果より, MTBD による SPAS-1 の微小管への結合は, SPAS-1 の安定な 6 量体形成を可能にし,その ATPase 活性を促進することが示唆された。一方,変異体を用いた解析から, SPAS-1 の微小管切断活性には,リング状オリゴマーの pore に位置する保存された芳香族残基とその周辺および pore 内部に位置する塩基性残基が重要であることを明らかにした(図 1 )。微小管の表面は,酸性残基が豊富な tubulin の C 末端がふらふらしている。表面プラズモン共鳴法を用いて, SPAS-1 が tubulin C 末端ペプチドと特異的に相互作用することを見出した。これらの結果から, SPAS-1 と微小管の相互作用は, MTBD による相互作用に加えて, AAA ドメインリングの pore 周辺と tubulin C 末端の相互作用も重要であることが判明し,図 2 に示すような spastin による微小管切断モデルを提唱した。これらの研究成果は, Genes Cells 誌電子版に先行発表された。
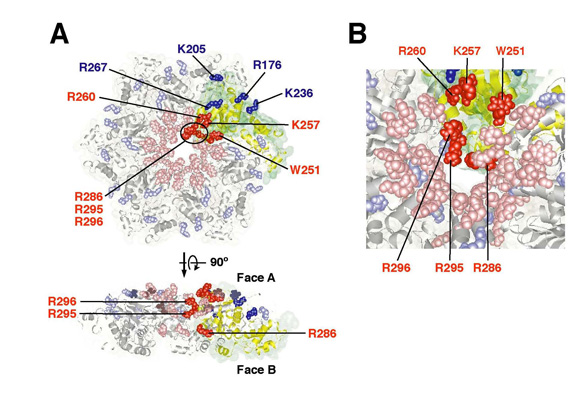
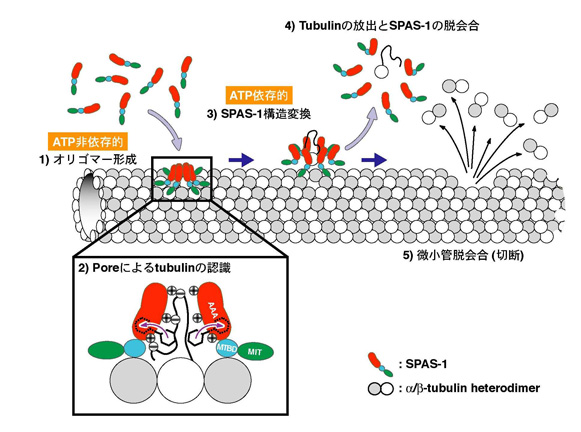
図 1 (上) SPAS-1 のリング状オリゴマーの pore 周辺に位置する塩基性残基は微小管の切断において重要である。 (A) SPAS-1 AAA ドメインの 6 量体リング構造のホモロジーモデリング。アラニンに置換することによって微小管切断活性が消失した残基を赤で,微小管切断活性には影響を及ぼさなかった残基を青で示す。数字はアミノ酸番号。 (B) Pore 入口と内部を拡大したものを示す。 Pore の入口に位置する W251 , R260 および R296 と内部に位置する R286 および R295 。
図 2 (下) Spastin による微小管切断モデル。 SPAS-1 は MTBD を介して微小管と結合し, ATP 非依存的に 6 量体を形成する。リング状オリゴマーの pore 周辺に位置する塩基性残基により tubulin C 末端を認識し, SPAS-1 の ATP 依存的な構造変化により微小管から tubulin が引っ張り出される。このステップが繰り返され,微小管脱会合が起こる。
