




Fujiwara Y., Horisawa-Takada Y., Inoue E., Tani N., Shibuya H., Fujimura S., Kariyazono R., Sakata T., Ohta K., Araki K., Okada Y., Ishiguro K. Meiotic cohesins mediate initial loading of HORMAD1 to the chromosomes and coordinate SC formation during meiotic prophase. PLOS Genetics 16(9): e1009048. (2020)
[ポイント]
・生殖細胞を作るために必要な減数分裂時に、染色体同士のマッチングを正常に行うための監視役を果たす因子が、コヒーシンとよばれるタンパク質の働きを借りて機能することを明らかにしました。
・父方由来、母方由来の染色体の遺伝情報の交換を行う場である軸構造の“大黒柱”として、コヒーシンが極めて重要な役割を担っていることを明らかにしました。
[研究の内容]
卵巣や精巣では減数分裂と呼ばれる特殊な細胞分裂が行われて卵子や精子が作り出されます。このとき父方、母方の2つの染色体が整列する「相同染色体の対合」とよばれる過程を経て減数分裂組換えによって遺伝情報の部分的な交換が行われます(図1)。「相同染色体の対合」は父方DNAと母方DNAとの間でDNA配列の交換を促進するのに必須の過程ですが、これがうまくいかないと減数分裂組換えが正常に起きずに卵子や精子の形成ができなくなってしまいます。この「相同染色体の対合」には正確に起きているかどうかを監視するメカニズムがあることが知られています。本研究では、精子・卵子の形成に必要な減数分裂の過程で父方由来、母方由来の染色体がマッチングして遺伝情報の交換を行う「相同染色体の対合」において監視役を果たす因子を呼び込む仕組みについて解析を行いました。
[成果]
先行研究では、HORMAD1と呼ばれるタンパク質が染色体の上に沿って出現し、相同染色体のマッチングがうまく行われているかを監視することが知られていました。本研究では減数分裂の際にHORMAD1がどのように染色体上に呼び込まれるのか、そのメカニズムについて検討しました。質量分析法を駆使した解析の結果、HORMAD1がSYCP2とコヒーシンとよばれる2つのタンパク質と結合することが判明しました。SYCP2とコヒーシンはいずれも減数分裂期の染色体を構成する主要な因子ですが、どちらがより重役を担っているかについてさらに検討したところ、ゲノム編集によりマウスのSYCP2遺伝子の働きをなくしてもHORMAD1がコヒーシンとの結合によって染色体上に現れて機能するのに対して、コヒーシンの働きをなくした場合にはHORMAD1が染色体上に正常に現れなくなることがわかりました。したがって「相同染色体の対合」に先駆けて、コヒーシンがHORMAD1を呼び寄せる目印として働いて染色体上に繋ぎ留める役割を担うことが明らかとなりました(図2)。減数分裂の過程で、染色体は「軸」と呼ばれる特殊な芯のような構造によって束ねられています。この染色体の「軸」構造は、父方と母方染色体のマッチングと遺伝情報の交換が正常に行われるための場として極めて重要な役割を担っていますが、コヒーシン分子が言わば染色体の軸構造の「大黒柱」として働くことがわかりました(図3)。
今回の成果はマウスを用いて検証されたものですが、ヒトの生殖細胞でも同じくコヒーシンやHORMAD1タンパク質が働いています。この仕組みがはたらかないと減数分裂組換え反応がうまく起こらなくなるため、精子・卵子形成が著しく低下し不妊となってしまいます。ヒトの不妊症は原因が不明とされる症例が多いことが知られています。今回の発見は、卵子・精子形成不全を示す不妊症の病態の解明に資することが期待されます
本研究は東京大学定量生命科学研究所の岡田由紀教授・藤原靖浩助教グループ、本学生命資源研究センターの荒木喜美教授グループ、発生研リエゾンラボとの共同によって行われました。本研究は文部科学省 科学研究費助成事業 新学術領域研究(非ゲノム情報複製)、文部科学省学術研究支援基盤形成・先端モデルプラットフォームおよび発生研共同利用の支援を受けて実施されました。
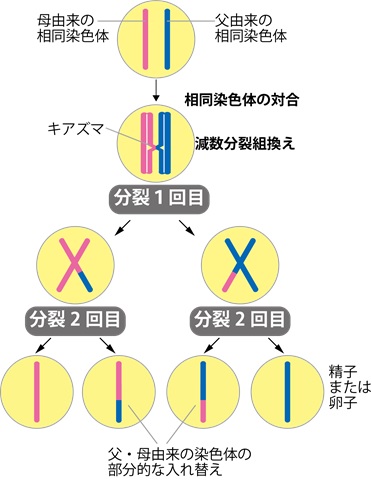
図1 減数分裂における父・母由来の相同染色体の対合と遺伝情報の部分的入れ替え

図2 相同染色体の対合の監視役HORMAD1の局在
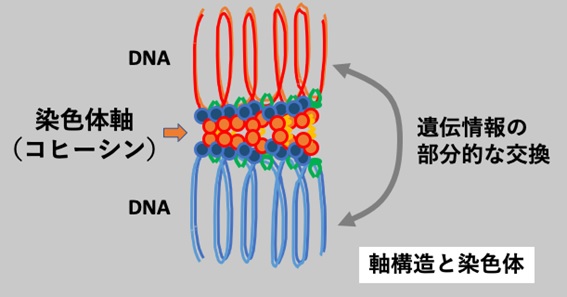
図3 相同染色体マッチングを監視するための染色体軸構造
