




Yuka Matsushita-Ishiodori, Kunitoshi Yamanaka, Hiroshi Hashimoto, Masatoshi Esaki, and Teru Ogura (2009) Conserved aromatic and basic amino acid residues in the pore region of C. elegans spastin play critical roles in microtubule-severing. Genes Cells Published Online: July 13 2009 .
Mutations of human spastin, an AAA (ATPases associated with diverse cellular activity) family protein, cause an autosomal dominant form of hereditary spastic paraplegia, which is characterized by weakness, spasticity and loss of the vibratory sense in the lower limbs. Recently, it has been reported that spastin displays microtubule-severing activity. We also previously reported that Caenorhabditis elegans spastin homologue SPAS-1 displays microtubule-severing (Matsushita-Ishiodori et al . (2007) Biochem. Biophys. Res. Commun . 359 , 157-162) . However, the detailed molecular mechanism of microtubule-severing remains unknown. Here we describe that SPAS-1 forms a stable hexamer in a concentration-dependent manner and that ATPase activity of SPAS-1 is greatly stimulated by microtubules. Furthermore, MTBD (microtubule binding domain) of SPAS-1 is essential for binding to microtubules. Taken these results together, we propose that MTBD of SPAS-1 plays a critical role in enrichment of SPAS-1 to microtubules, where SPAS-1 is concentrated and able to form a stable hexamer, subsequently its ATPase activity is stimulate d. On the other hand, our mutational analyses revealed that the conserved aromatic and basic amino acid residues in the pore region are important for microtubule-severing (Fig. 1). We also detected the direct interaction of the extremely acidic C-terminal polypeptide of tubulin with SPAS-1. Furthermore, our results imply that there are at least two recognition sites in the pore region, one is a set of basic amino acid residues on Face A and the other is a set of basic amino acid residues inside the pore. Our results also indicate that the recognition on Face A is preceded to the recognition of inside the pore, that is, substrates may be translocated from Face A side to inside the pore. Consequently, we propose a new model of microtubule-severing by SPAS-1 (Fig. 2).
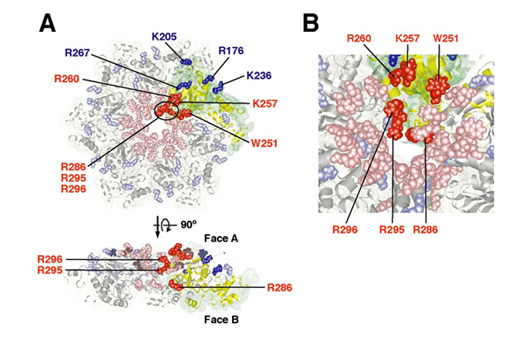
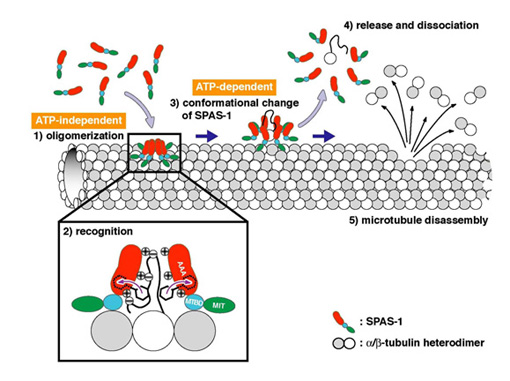
Fig. 1 Requirement of solvent-exposed, basic amino acid residues in the pore region of SPAS-1 for microtubule-severing. (A) Mutated residues are shown on the modeled hexameric structure of SPAS-1. In the case that mutant proteins lost microtubule-severing activity, the amino acid residues are shown in red. In the case that mutant proteins still possessed it, they are in blue. (B) The detailed view highlights the entrance of the pore and inside the pore.
Fig. 2 A model for microtubule-severing by spastin. SPAS-1 assembles into a hexamer on microtubules. This step requires MTBD. By ATP-dependent conformational changes SPAS-1 oligomers give rise to disintegration of tubulin from microtubules and release of tubulin together with SPAS-1 oligomers. Subsequently , SPAS-1 oligomers are dissociated due to the disappearance of anchorage. These tubulin extraction steps are repeated and leading to microtubule disassembly.
