Establishment of hTSCs (Okae et al. Cell Stem Cell 2018)
In 1998, Dr. Satoshi Tanaka and his colleagues succeeded in the establishment of mouse TSCs. Since then, many researchers have tried to establish hTSCs but only failed. To understand how human trophoblast cells proliferate in vivo, we isolated trophoblast cells from human placentas and analyzed their gene expression patterns. This analysis revealed that Wnt and EGF signaling pathways may be essential for the proliferation of human trophoblast cells. Based on this finding, we optimized the culture conditions and succeeded in the establishment of hTSCs.
hTSCs can be maintained in vitro almost indefinitely without losing their ability to differentiate into two trophoblast lineages, syncytiotrophoblast and extravillous cytotrophoblast (Fig. 1).
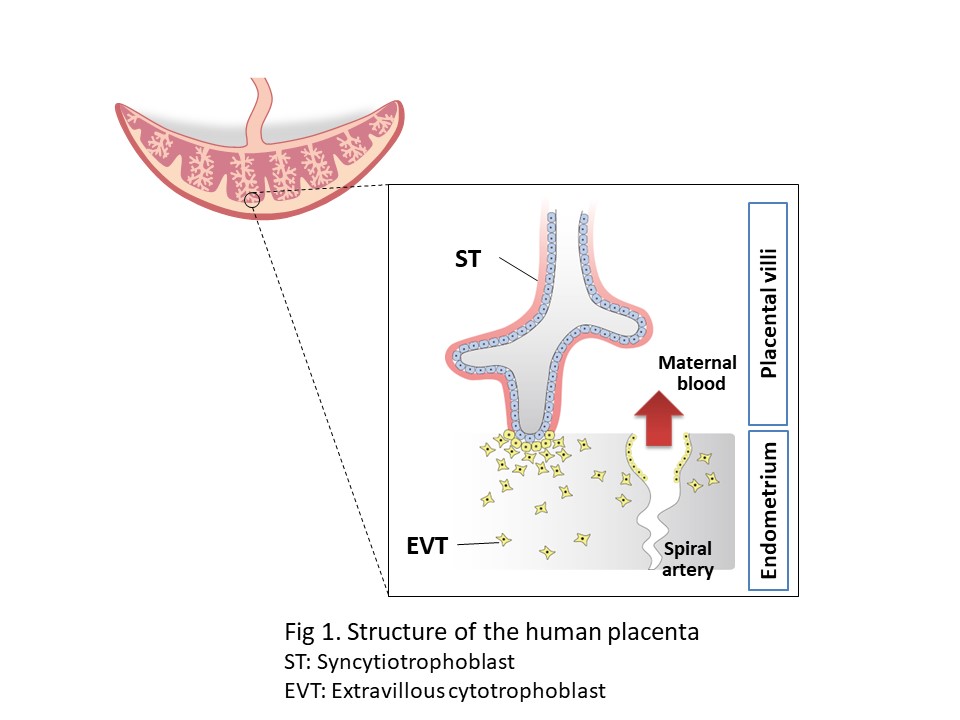
Syncytiotrophoblast cells mediate the exchange of gasses and nutrients between the fetus and the mother, as well as producing hormones to maintain pregnancy. Extravillous cytotrophoblast cells invade the endometrial tissue and remodel the spiral artery, which may play an important role in controlling maternal blood flow.
Gene expression analyses suggest that hTSCs are most similar to peri-implantation trophoblast cells. Consistently, hTSCs injected into immunodeficient mice differentiated into primitive syncytiotrophoblast cells, specialized cells critical for implantation, and infiltrated surrounding mouse tissue (Fig. 2). We also detected large amounts of pregnancy hormones in the blood of the recipient mice. These data suggest that hTSCs can reproduce key features of human implantation.
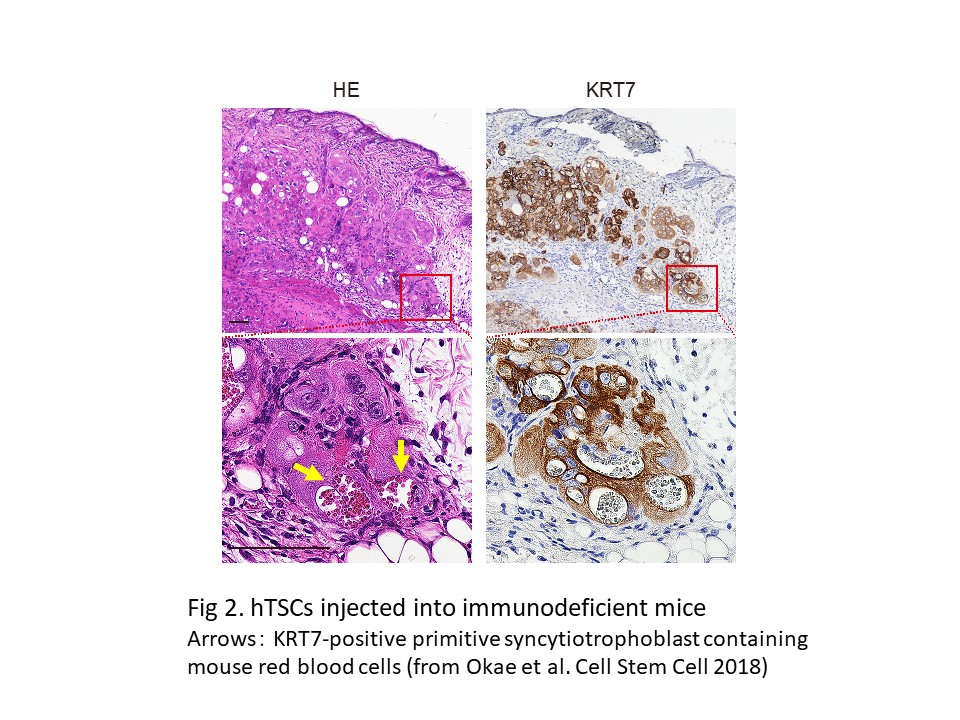
Based on these findings, we conclude that hTSCs provide a useful model for understanding human placental development and implantation.
Biomedical applications of hTSCs (Takahiashi et al. PNAS 2019, Kobayashi et al. Nature Commun 2022)
Using the culture technique of hTSCs, we have reported the utility of hTSCs in studies of pregnancy complications and early human development. We would like to introduce our two recent studies.
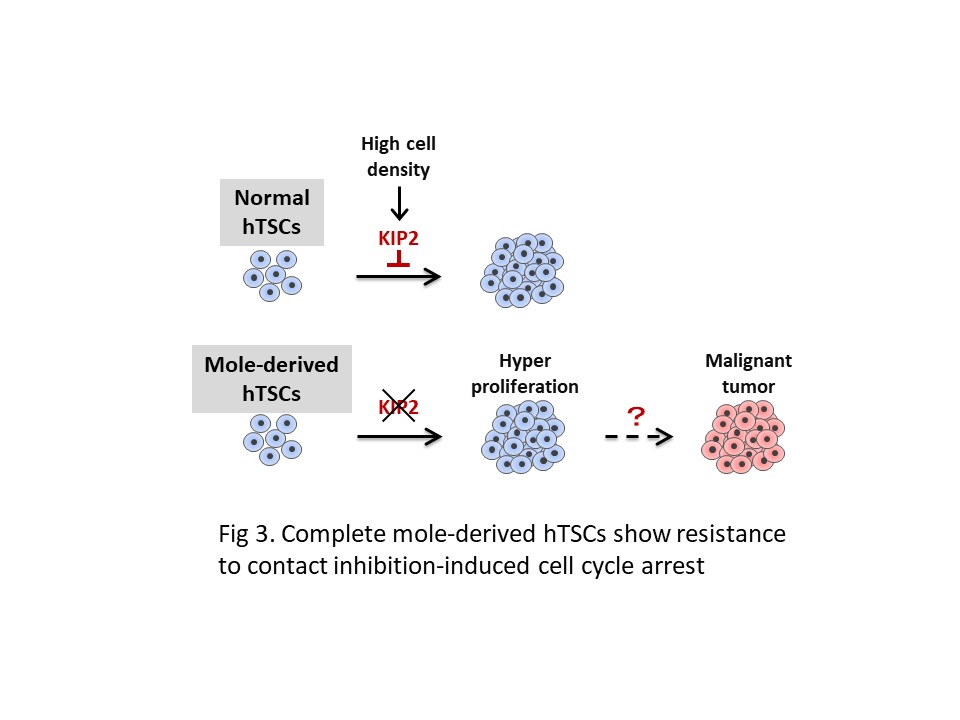
The first is a study of complete mole. Complete mole originates from androgenetic embryos that have only the paternal genome and is characterized by abnormal proliferation of trophoblast cells. In this study, we derived hTSCs from complete mole tissues and analyzed their characteristics in detail. We found that hTSCs derived from complete mole show resistance to contact inhibition-induced cell cycle arrest. Moreover, we revealed that p57/KIP2, a negative regulator of cell cycle, is not expressed in complete mole-derived hTSCs, which confers resistance to contact inhibition (Fig. 3). These data highlight the utility of hTSCs in the study of pregnancy complications.
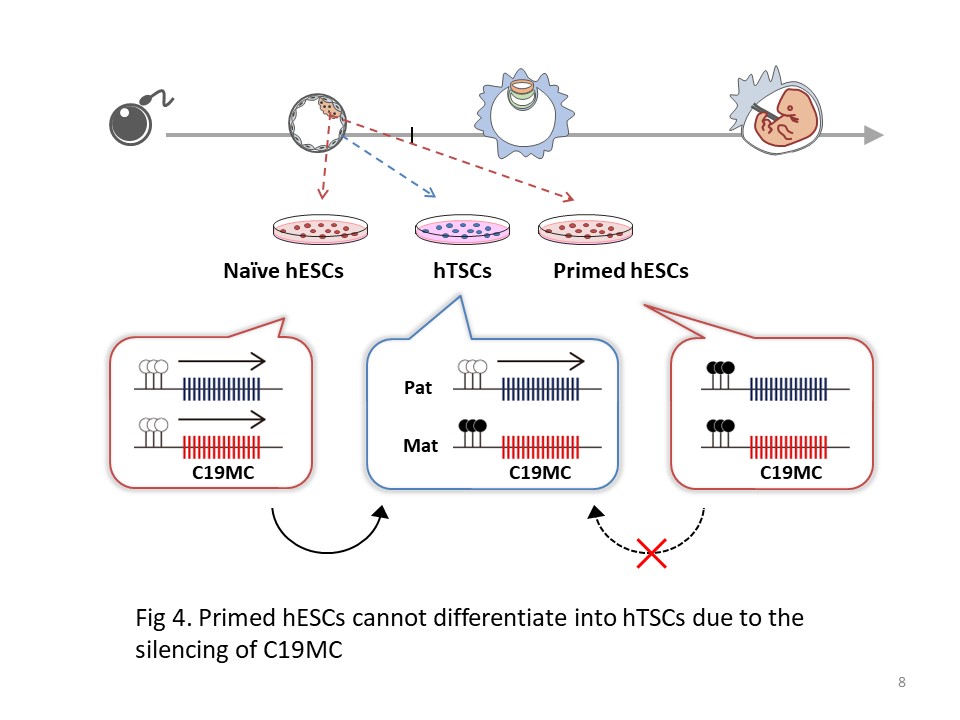
Another study is the establishment of a conversion system from hESCs to hTSCs. By comparing naive and primed hESCs, which represent pre- and post-implantation epiblast cells respectively, we found that naive hESCs can differentiate into hTSCs, but primed hESCs cannot. We also utilized genome and epigenome editing techniques and revealed that the DNA methylation status of the primate-specific miRNA cluster C19MC explains the phenotypic differences between naive and primed hESCs. From these data, we suggest that C19MC may function as an epigenetic barrier preventing transdifferentiation from epiblast cells into trophoblast cells after implantation (Fig. 4).