









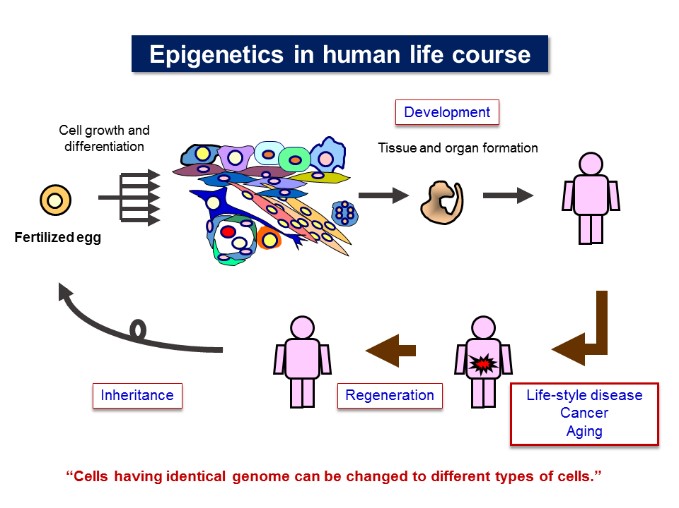
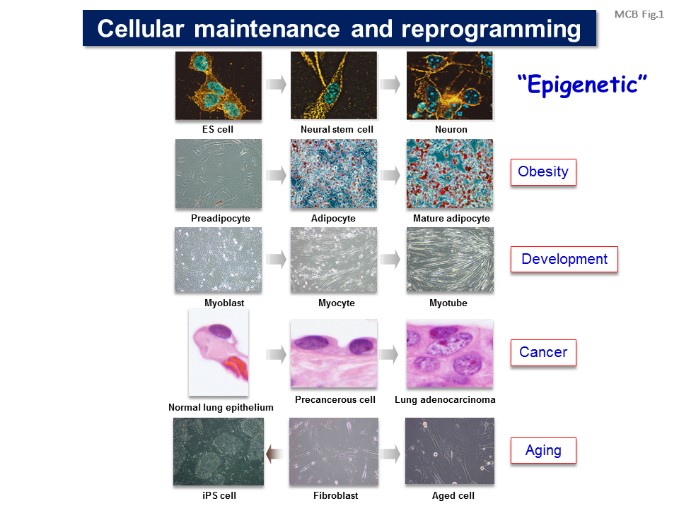
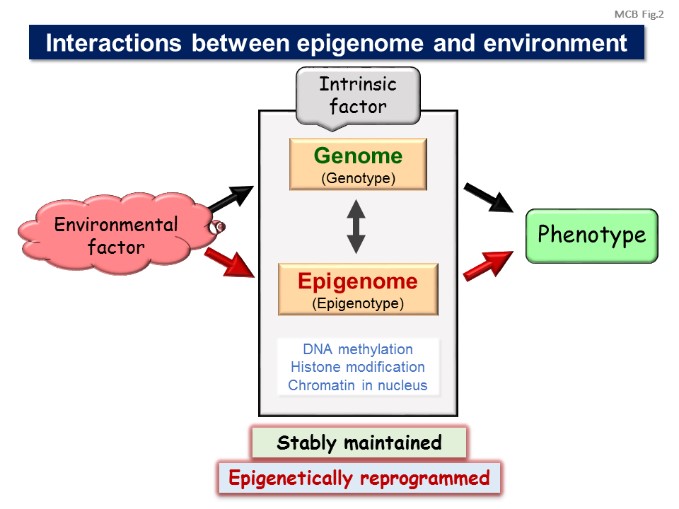
Our laboratory is studying the molecular basis of epigenetic cell regulation in development and human diseases. The term epigenetic is defined as “heritable changes in gene expression that occur without a change in DNA sequence”. Epigenetic regulation may include cytosine methylation, histone modification, chromatin formation, and nuclear structure.
We are studying how these epigenetic factors control gene expression and cellular function; 1) investigating the molecular basis of epigenome and gene control; 2) studying the epigenetics of energy metabolism; 3) identifying the mechanism involving in epigenetics of cancer and inflammation; 4) studying epigenome of cellular development and senescence; 5) detecting nuclear structure, function and dynamics; and 6) testing epigenetic technology useful for medical diagnosis and therapy. [Figure 1]
【Figure 1】
1. Molecular basis of epigenome and gene control
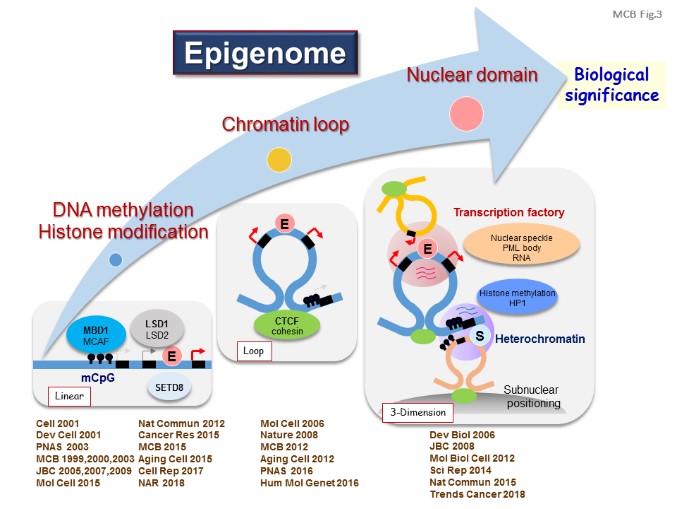
Methylated DNA is specifically recognized by a set of proteins called methylated DNA-binding proteins, which belong to three different families in mammals: the MBD proteins, the zinc finger (Kaiso) proteins, and the SRA (UHRF) domain proteins. Once bound to methylated DNA, these proteins translate the DNA methylation signal into appropriate functional states, through interactions with various partners.
We investigated the function of MBD1 and found MBD1-containing chromatin-associated factor (MCAF), also known as ATFa-associated modulator (AM) and activating transcription factor 7-interacting protein (ATF7IP), which mediates gene regulation (Cell, 2001; MCB, 2003).
Long-range regulatory elements and higher-order chromatin structure coordinate the expression of multiple genes in cluster, and CTCF/cohesin-mediated chromatin insulator may be a key in this regulation, together with CTCF/CHD8 chromatin remodeling (Mol Cell, 2006; Nature, 2008). The human apolipoprotein (APO) A1/C3/A4/A5 gene region, whose alterations increase the risk of dyslipidemia and atherosclerosis, is partitioned at least by three CTCF-enriched sites and three cohesin protein RAD21-enriched sites, resulting in formation of two transcribed chromatin loops by interactions between insulators (EMBO J, 2009). [Figure 2, 3]
【Figure 2】
【Figure 3】
2. Epigenetics of energy metabolism
Environmental factors such as nutritional state may act on the epigenome which consequently contributes to the metabolic adaptation of cells and the organisms. The lysine demethylase LSD1 is a unique nuclear protein that utilizes flavin adenine dinucleotide (FAD) as a cofactor. We showed that LSD1 epigenetically regulates energy expenditure genes in adipocytes depending on the cellular FAD availability (Nat Commun, 2012). We also revealed that LSD1 is an integrative regulator of aerobic glycolysis in cancer (Cancer Res, 2015; Int J Cancer, 2016), and is a determinant of metabolism-fiber type switch in muscle cells (Nucleic Acids Res, 2018). In addition, we reported that another FAD-dependent demethylase, LSD2, protects hepatic cells from lipotoxicity by repressing lipid transport genes (MCB, 2015). Our data shed light on an essential mechanism of energy utilization which might explain how cells determine their energy strategy depending on nutritional availability. [Figure 4, 5]
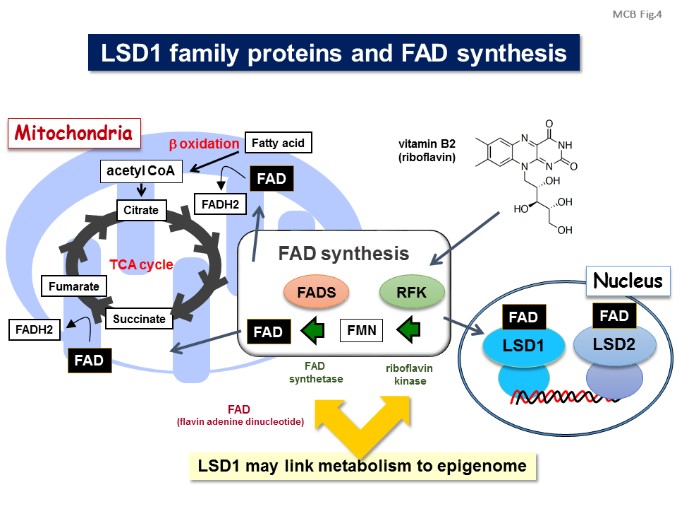
【Figure 4】
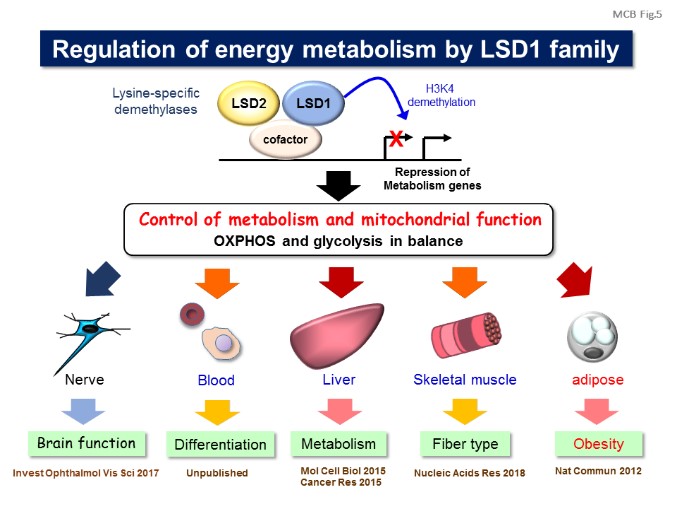
【Figure 5】
3. Epigenetics of cancer and inflammation
Estrogen receptor-α (ER)-positive breast cancer cells undergo hormone-independent proliferation after deprivation of estrogen, leading to endocrine therapy resistance. Up-regulation of the ER gene (ESR1) is critical for this process, but the underlying mechanisms remain unclear. Recently, the combination of transcriptome and fluorescence in situ hybridization analyses revealed that estrogen deprivation induced a cluster of non-coding RNAs that defined a large chromatin domain containing the ESR1 locus. We named these RNAs as Eleanors (ESR1 locus enhancing and activating non-coding RNAs) (Nat Commun, 2012).
Eleanors were present in ER-positive breast cancer tissues and localized at the transcriptionally active ESR1 locus to form characteristic RNA foci. Depletion of one of Eleanors, upstream (u)-Eleanor, impaired cell growth and transcription of intragenic Eleanors and ESR1 mRNA, indicating that Eleanors cis-activate the ESR1 gene. Furthermore, resveratrol, a kind of polyphenol, was found to repress these RNAs and inhibit the proliferative activity of breast cancer cells which had acquired estrogen-independent growth. Thus, Eleanors-mediated gene activation plays an essential role in the adaptation of breast cancer cells. We clinically discuss diagnostic and therapeutic targets for endocrine therapy-resistant breast cancer (Trends Cancer, 2018).
We further investigated the effects of tumor necrosis factor (TNF) signaling on spatiotemporal enhancer-promoter interactions in the human tumor necrosis factor (TNF)/lymphotoxin (LT) gene locus, mediated by CTCF-dependent chromatin insulators (MCB, 2012). The cytokine genes LTα, TNF, and LTβ are differentially regulated by NF-κB signaling in inflammatory and oncogenic responses. We identified at least four CTCF-enriched sites with enhancer-blocking activities and a TNF-responsive TE2 enhancer in the TNF/LT locus. The insulators mediate the spatiotemporal control of enhancer-promoter associations in the TNF/LT gene cluster.
In addition, we are investigating the epigenetics of inflammation memory. Immunological memory in vertebrates is generally thought to be controlled by memory T and B cells. However, recent studies in invertebrates including mosquito and drosophila that lack adaptive immune systems can also exhibit inflammation memory, which means second infection is exclusively affected by first infection without lymphocytes (T and B cells). Inflammation memory is also reported in vertebrates such as adaptive immune system-deficient Rag1 knockout mice. However, it is still unknown how initial inflammation affects second infection and how inflammation memory is maintained. We thus investigate whether epigenetic changes by first inflammation contributes to the innate immune responses against second inflammation with respect to various epigenetic status including DNA methylation, histone methylation, histone acetylation, chromatin loop, nuclear compartments including euchromatin and heterochromatin. [Figure 6, 7]
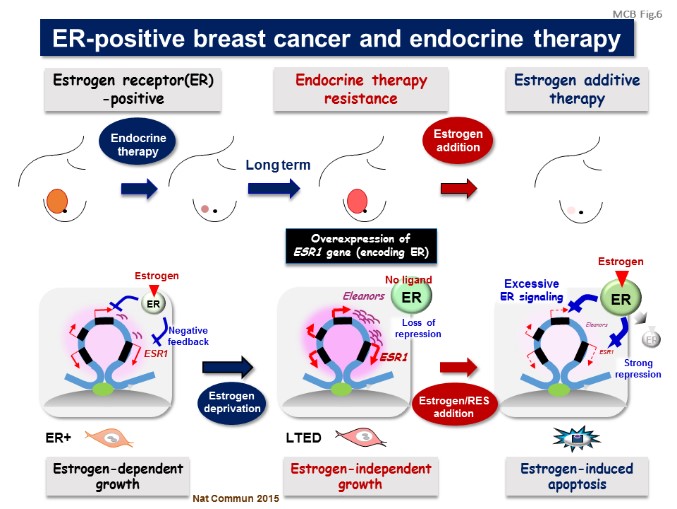
【Figure 6】
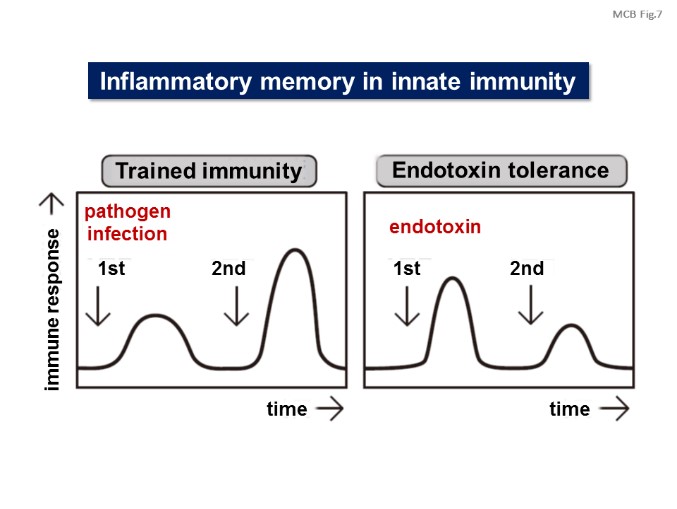
【Figure 7】
4. Epigenetics of development and senescence
The INK4/ARF locus encodes p15INK4b, ARF, and p16INK4a genes in human chromosome 9p21, the products of which are known as common key reprogramming regulators. Compared with growing fibroblasts, CTCF is remarkably up-regulated in iPS cells with silencing of the three genes in the locus and is reversely down-regulated in senescent cells with high expression of p15INK4b and p16INK4a genes. There are at least three CTCF-enriched sites in the INK4/ARF locus, which possess chromatin loop-forming activities. These results suggest that senescent cells have distinct higher-order chromatin signature in the INK4/ARF locus (Aging Cell, 2012).
We revealed that metabolic reprogramming is essential for the progression of cellular senescence, and that histone methyltransferase SETD8/PR-Set7 prevents cellular senescence and senescence-associated metabolic remodeling through regulating specific gene expression (Aging Cell, 2015; Cell Rep, 2017). [Figure 8, 9]
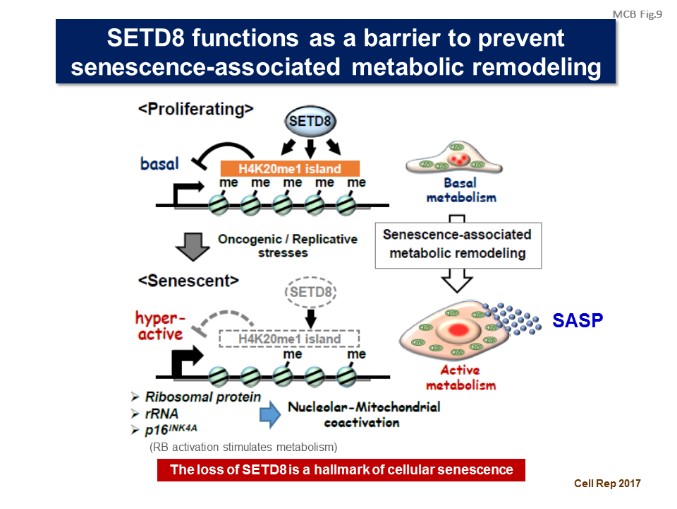
【Figure 8】
【Figure 9】
5.Nuclear structure, function and dynamics
The nucleus is the origin of cellular function, because it can govern biological information within it. Transcription, RNA dynamics, DNA replication, DNA damage responses, and recombination can be regulated by accumulation of key molecules and their complexes at the respective unique domains in the nucleus. The nuclear domains are actively formed and dispersed in response to the cell environments. During the cell division, the nuclear architectures and domains are broken up and then re-established. Because many of the etiologies in cancer, autoimmune diseases and neurological disorders target the components of the nucleus, we investigated the dynamic structure and function of the nucleus, from the physiological and pathological aspects (JBC, 2008; Mol Biol Cell, 2012). [Figure 10, 11]
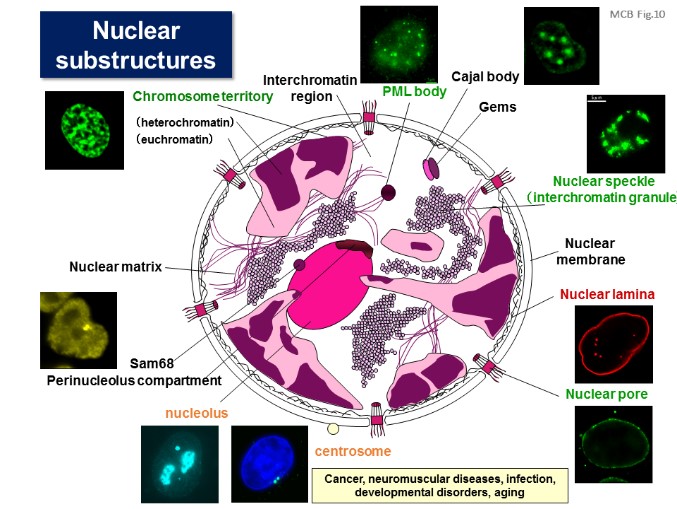
【Figure 10】
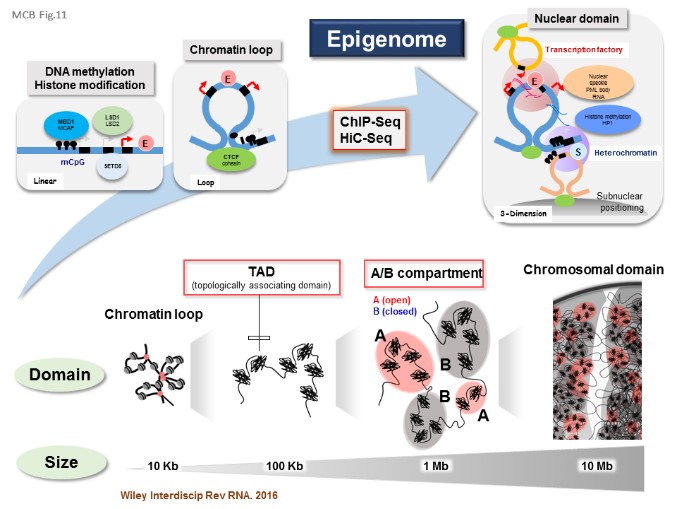
【Figure 11】
6.Epigenetic technology useful for medical diagnosis and therapy
Based on our recent progresses, MCAF1 can be used for diagnostic and therapeutic agents in human cancers, since this protein is highly expressed in a variety of cancers.
We then found that the loss of LSD1 function either by siRNAs or by selective inhibitors in adipocytes induces a number of regulators of energy expenditure and mitochondrial metabolism resulting in the activation of mitochondrial respiration. In the adipose tissues from high fat diet-fed mice, expression of LSD1-target genes was reduced compared with that in normal-diet mice, which can be reverted by suppressing LSD1 function.
Further, we established methods to quantitatively assess the imaging data of nuclear domains or substructures such as PML bodies and nucleolus using the pattern recognition software wndchrm (weighted neighbour distances using a compound hierarchy of algorithms representing morphology) (Sci Rep, 2014). Collectively, we are investigating epigenetics from the viewpoint of an interaction between gene and environment.[Figure 12]
【Figure 12】









