



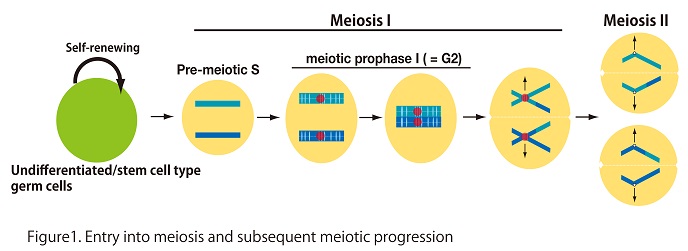
The meiotic cell cycle consists of a single DNA replication followed by two rounds of chromosome segregation (meiosis I and meiosis II), which halves the chromosome number in haploid gametes. The structure and behavior of chromosomes during meiosis are remarkably different to that in mitosis.
Meiosis is a special cell cycle that is a modification of the canonical mitotic cell cycle. Understanding the mechanisms how the meiotic cell cycle is regulated for correct assembly of specialized chromosome structure and timely order of chromosomal events is important because dysregulation of meiosis often leads to infertility and pregnancy losses. Our laboratory is investigating the molecular mechanisms of meiosis from the following three aspects: (1) molecular mechanisms of induction of meiosis from a mitotic state in precursor germ cells, (2) molecular mechanisms of cell cycle regulation that provide a crucial difference between meiosis and mitosis, and (3) the molecular basis of a meiosis-specific chromosomal structure that ensures unique chromosomal events during meiosis.
(1) Investigating the molecular mechanisms of meiosis induction
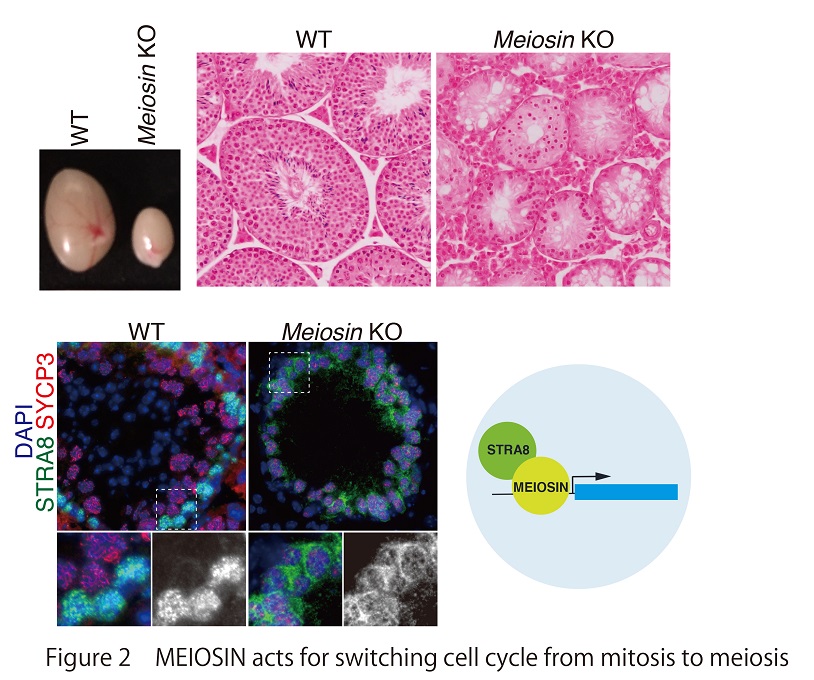
In mammals, undifferentiated/stem cell type germ cells (primordial germ cell in female or spermatogonia in male) have a potential to enter meiosis. After mitotic growth, a subpopulation of such germ cells spontaneously progress to meiotic S-phase expressing STRA8, which is triggered by external signal of retinoic acid (Fig1). Importantly, it is assumed that induction of meiosis accompanies switching the cell cycle regulation from “mitotic type” to “meiotic type”. Thus, the regulatory mechanism of meiotic initiation is long-standing enigma in mammals.
It has been known that the induction of STRA8 by the retinoic acid (RA) signaling has a pivotal role during meiotic initiation. STRA8, which is expressed in response to RA, is thought to be a key factor promoting meiotic initiation. However, how STRA8 is involved in meiotic initiation has remained elusive. Recently, our group identified MEIOSIN (Meiosis initiator) as a germ cell-specific factor, which associates with STRA8 (Dev Cell 2020) (Fig2). MEIOSIN, like STRA8, is expressed in response to RA and plays an essential role in meiotic initiation both in male and female. MEIOSIN together with STRA8 acts as a crucial transcription factor to drive meiotic gene activation. Thus, a key finding of our study is that MEIOSIN collaborating with STRA8 plays a central role in germ cell fate decision from mitosis to meiosis in mammals.
Our current aim is to further investigate the mechanisms of meiotic initiation. Since we identified many uncharacterized genes that are targeted by the MEIOSIN-STRA8 complex, functional analyses of such downstream genes are the current focus of our project.
(2) Studying the molecular mechanisms of cell cycle regulation in meiosis
Our specific aim in this project is to elucidate molecular differences in cell cycle regulation between mitosis and meiosis. In germ cells, meiosis-specific events are programed in the context of cell cycle regulation (Fig. 1). Notably, meiotic prophase I, which is equivalent to the G2 phase in the somatic cell cycle, is prolonged so that meiosis-specific chromosomal events progress sequentially. Moreover, one round of S-phase is missing between the first (M I) and second M phases (M II), which is in sharp contrast to the somatic cell cycle. Thus, it is assumed that canonical cell cycle regulation is diversified and modified in meiosis. However, the underlying mechanism of the meiotic cell cycle is still largely elusive.
As it is well known that chromosomal events, such as DNA replication, recombination, repair and chromosome segregation, are driven by cell cycle regulation in somatic cells, it is also important to consider the mechanisms of meiotic chromosome dynamics with respect to the cell cycle. Thus, our current approach is to identify meiosis-specific substrates and regulatory subunits for E3 ubiquitin ligases in germ cells that are crucial for cell cycle progression.
(3) Studying the molecular basis of a meiosis-specific chromosomal structure
When chromosomes are replicated in the S-phase, sister chromatids are held together by a mechanism (sister chromatid cohesion) which enables accurate chromosome segregation in both mitosis and meiosis. Sister chromatid cohesion is mediated by an evolutionary conserved multi-protein complex, cohesin. Moreover, cohesin acts for transcriptional regulation by collaborating with insulator binding factors in somatic cells. Importantly, there are two types of cohesins: mitotic-type and meiotic-type (Fig. 2).
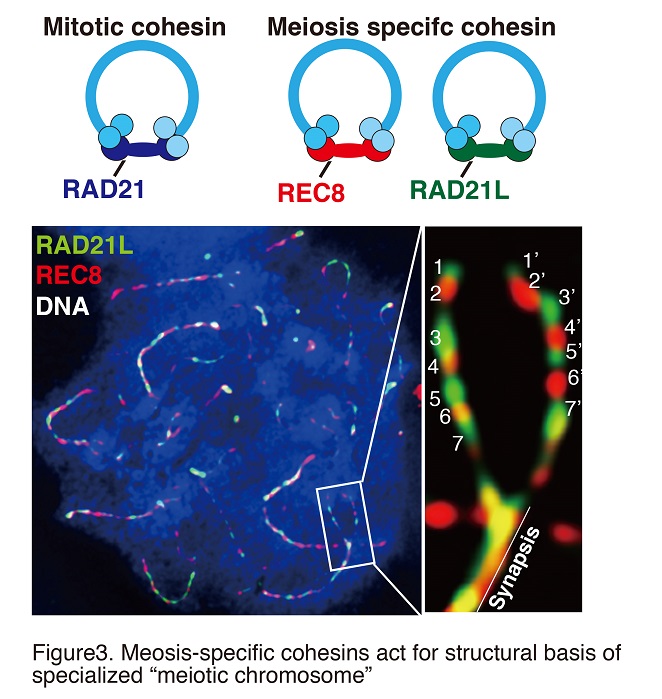
During meiotic prophase I, sister chromatids are organized into proteinaceous structures called chromosome axes on which the synaptonemal complex is assembled. Consequently, meiotic cohesin acts as a structural basis for chromosomes (Fig. 3). Homologous chromosomes then undergo pairing, synapsis, and meiotic recombination yielding a physical linkage between homologs called chiasmata (Fig. 4). Meiosis-specific cohesin plays a crucial role in all of these sequential chromosomal events during meiosis. In mammalian meiotic cells, two different types of cohesin complexes exist which contain either subunit REC8 or subunit RAD21L. The REC8 and RAD21L subunits determine the spatiotemporal distribution patterns of distinct cohesin complexes on chromosomes during meiosis (Fig. 2). Thus, REC8- and RAD21L-cohesins play different roles in chromosomal dynamics during meiosis.
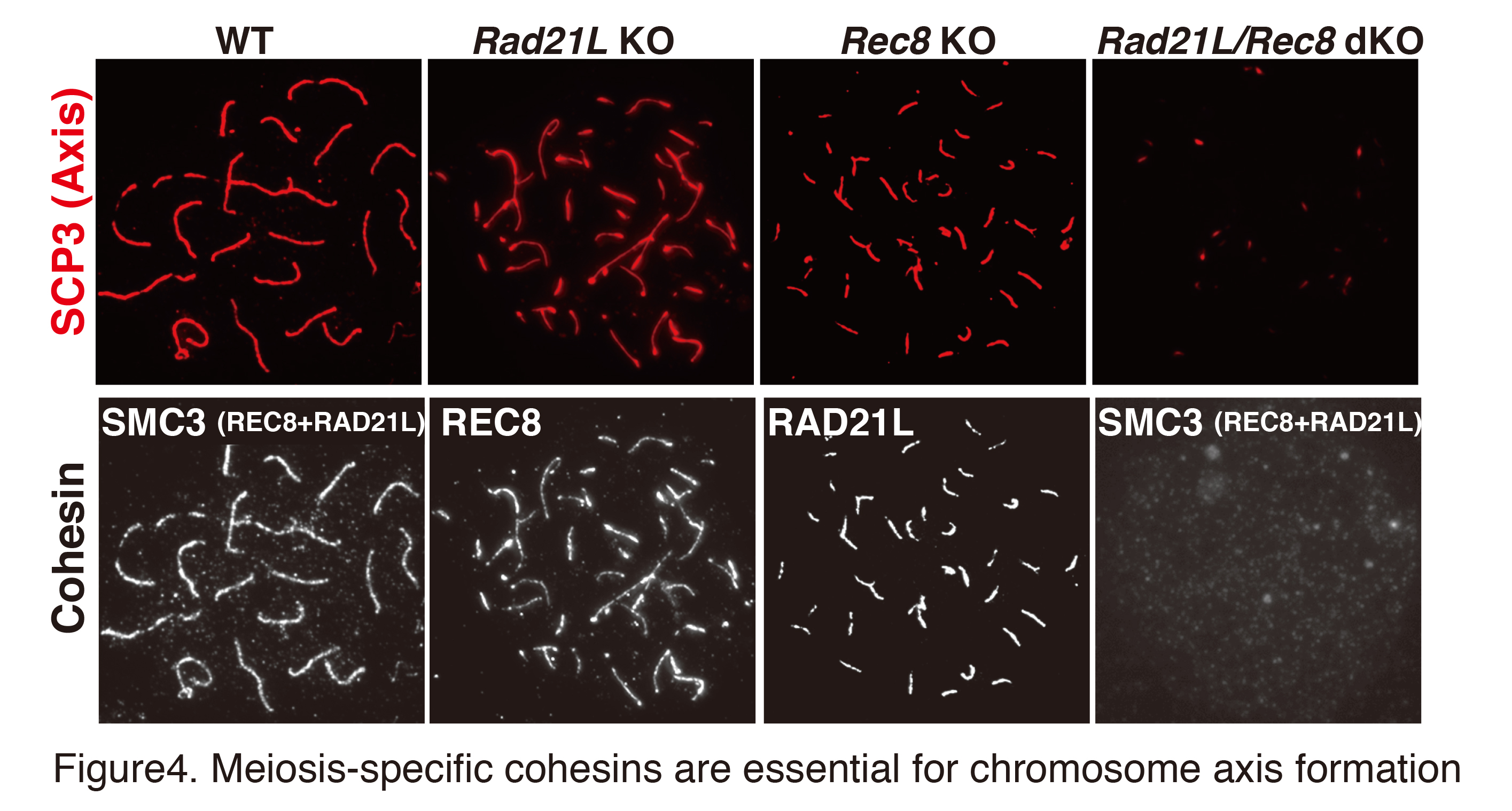
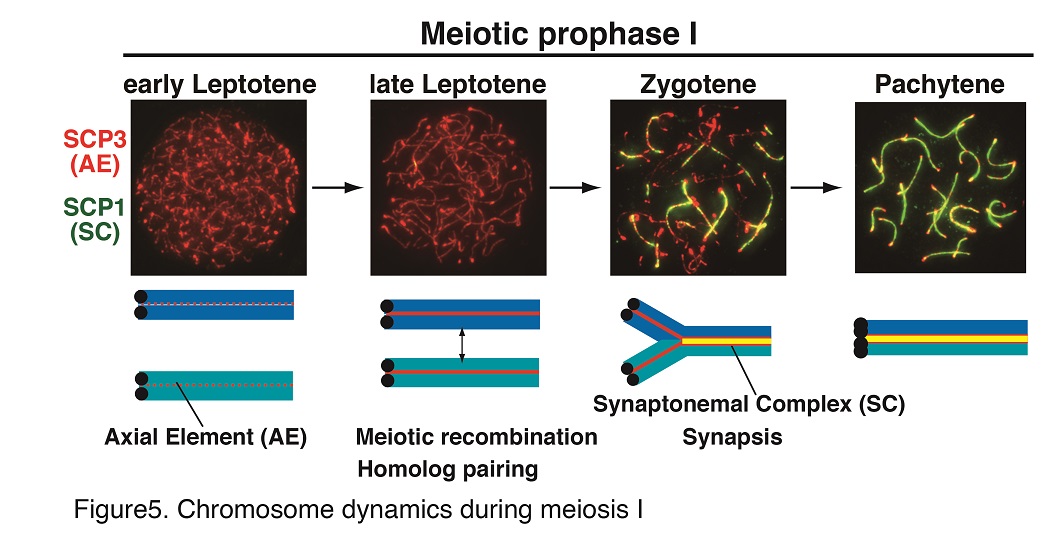
Notably, REC8- and RAD21L-cohesins do not have the ability to localize on chromosomes when they are expressed in somatic cells, suggesting that some factor may be missing for loading meiotic cohesins on somatic chromosomes. Our specific aim in this project is to elucidate the molecular mechanisms that determine how chromosome loading of distinct meiotic cohesins are licensed during meiosis. Since the pre-meiotic S-phase is the earliest time point when meiotic cohesins are detected, this project will be focused on identifying a putative cohesion loader during this period which would lead to reorganization of meiotic chromosomes.




