









Ai Johjima, Kentaro Noi, Shingo Nishikori, Hirotsugu Ogi, Masatoshi Esaki, Teru Ogura. Microtubule severing by katanin p60 AAA+ ATPase requires the C-terminal acidic tails of both α- and β-tubulins and basic amino acid residues in the AAA+ ring pore. J. Biol. Chem. 290: 11762-11770 (2015)
微小管は主にα-tubulinとβ-tubulinから構成される管状の高分子重合体である。微小管は,末端でtubulin分子が重合や脱重合をすることによって伸長・短縮し,細胞分裂の有糸分裂期での紡錘体の形成と染色体分配,細胞骨格として細胞の形態維持,モータータンパク質のレールとして細胞内輸送などさまざまな細胞機能に重要な役割を果たしている。Katanin p60は,ATP加水分解のエネルギーを利用して基質タンパク質の構造を変換するAAA (ATPases associated with diverse cellular activities)タンパク質の一つで,微小管構造を中央から切断する酵素であるが,その分子機構の詳細は明らかになっていない。
分子細胞制御分野(小椋 光教授)の城島 愛研究員らは,katanin p60による微小管切断の分子機構の一端を明らかにした。AAAタンパク質は一般に6量体のリング構造を形成し,中央の孔に突き出たループ構造(poreループ)には芳香族アミノ酸残基が保存されている。さらにkatanin p60の場合には,その芳香族アミノ酸残基の前後に塩基性アミノ酸残基も保存されている。本研究では,katanin p60のporeループにあるチロシン残基およびその前後の塩基性残基(リジンおよびヒスチジン残基)をそれぞれアラニンに置換した部位特異的変異体を作製した。いずれの変異体もATPase活性は保持していたが,微小管切断活性は示さなかった。したがって,poreループに保存された塩基性残基および芳香族残基は微小管切断に必須であることが明らかとなった。
一方,微小管表面にはα-tubulinおよびβ-tubulinのC末端領域が露出しており,この領域は酸性残基に富み,特定の構造をとらないdisorder領域であると考えられている。そこで,遺伝子操作によってα-tubulinおよびβ-tubulinのいずれか,または両方のC末端disorder領域を持たない改変微小管を作製し,微小管切断活性を測定した。その結果,katanin p60はいずれの改変微小管と相互作用するが,切断できないことを発見した。したがって,katanin p60による微小管切断には,α-tubulinおよびβ-tubulin両方のC末端disorder領域が必要であることが明らかとなった。
これらの解析により,katanin p60は微小管切断の際,両tubulinのC末端disorder領域を同時にリング孔に引き込み,tubulinの一部分をアンフォールドすることによってtubulin全体の構造変化を誘発し,それによってtubulin 2量体を引き抜き,微小管構造を破壊するという新規モデルを提唱した(図)。この研究成果は,2015年5月Journal of Biological Chemistry 誌に掲載された。
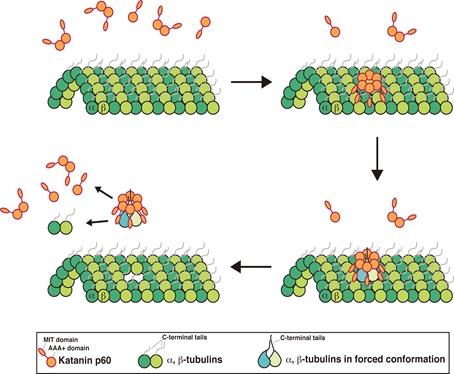
図 Katanin p60による微小管切断機構モデル図
Katanin p60は微小管上でオリゴマーを形成し,N末端にあるMIT(Microtubule interacting and trafficking)ドメインで微小管と相互作用する。両tubulin C末端disorder領域をkatanin p60のporeループの孔に引き込み,tubulinの一部をアンフォールドすることによってtubulin全体の構造変化を引き起こし,tubulin分子を微小管から引き抜くことができる。これを繰り返すことで微小管の脱重合を促進し,微小管構造を破壊する。


