Introduction of Our Research
Our body consists of a wide variety of cells. For example, an adult human body is composed of about 60 trillion, more than 200 kinds of cells. These cells can be classified into two types: somatic and germ cells. Somatic cells such as muscle, neuron, and skin cells make up the majority of our body and are essential for an individual to survive. At the end of individual’s life, the somatic cells accomplish all cellular functions and are destined to die. In contrast, although germ cells are dispensable for an individual to survive, they are the only cell types capable of transmitting genetic information from parents to children and creating a life for the next generation. Furthermore, all the cells comprising our body are produced from only one fertilized egg, which has been formed by joining of sperm and egg. Therefore, the germ cells can be regarded as the “ultimate” stem cells.
In many animal species, there is a specific cytoplasmic region within eggs, called the germ plasm. The cells inheriting the germ plasm become germ cells. It has been known that specific mRNAs and proteins are localized in the germ plasm. Unfortunately, the means by which the germ plasm is formed and maintained, and regulates germ cell determination remain largely unknown.
Our research team aims to clarify the mechanism of germ cell formation usingDrosophila as a model organism. Drosophila , fruit fly, is a small insect with 2-3 mm in length, and can be found anywhere in the world. From an evolutionary viewpoint,Drosophila is the organism that is phylogenetically very distant from human. On the contrary, from genomic and genetic viewpoints, Drosophila and human use similar cellular regulatory systems and have a very close relationship. We therefore believe that the knowledge gained from Drosophila research will greatly contribute to human health science and medical science research.
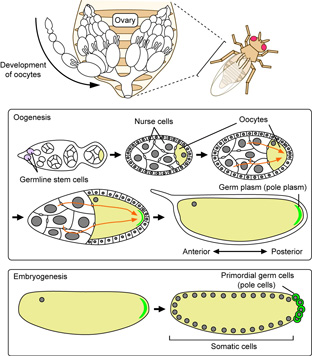
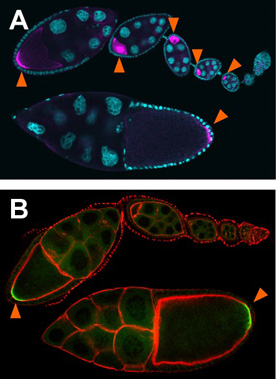
A process where Drosophila eggs are formed is called “oogenesis”. In the course of this process, mRNAs that are required to form the germ plasm are transported to the posterior pole of the oocyte (shown in pink on Panel A). The only mRNAs that have reached at the posterior pole of the oocyte are able to produce protein (shown in green on Panel B). We intend to elucidate the mechanisms by which mRNA localization and translational control are coupled, including the means of how mRNAs that encode proteins essential for the formation of the germ plasm is translationally repressed until they are localized to the oocyte posterior.
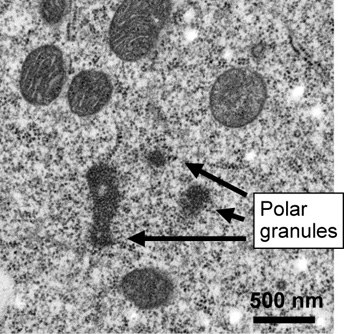
When examining the germ plasm in the Drosphila egg under electron microscopy, we can observe a specialized structure called “polar granules (germ granules)”. The polar granules are small organelles with their diameter of up to 500 nm (1/2000 mm), however, they contain specific RNAs and proteins which play essential role in germ cell formation. We are studying how these RNAs and proteins are aggregated to assemble germ granules ( dynamics of RNP complex ).
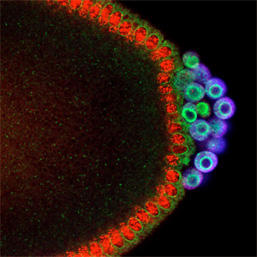
The cells inheriting germ plasm become pole cells, primordial germ cells, in this species. During the period when pole cells are formed in the embryo, numerous signals promoting somatic cell differentiation are traveling throughout the embryo including pole cells. These signals will possibly force pole cells to differentiate into somatic cells. To avoid this impact, pole cells have established a mechanism that actively inhibits transcription of genes associated with differentiation into somatic cell so that pole cells can maintain germ cell characteristics. Our research team is trying to unveil this mechanism.